Гидроусилитель руля, схема и принцип работы
На заре автомобилестроения никто, надо полагать, и думать не думал о том, чтобы с помощью какого-либо приспособления помогать водителю крутить баранку. Машины были легкими, колеса узкими, да и скорости небольшие. У первых автомобилей и руля в привычном понимании даже не было. Так – рычажок трамвайный.
Однако с появлением грузовиков большой грузоподъемности труд шофёра становился всё более тяжелым, крутить становящееся всё более тугим рулевое колесо несколько часов кряду было уже просто утомительно. Так появился гидроусилитель руля – изобретение, которое, как множество ему подобных, отчасти своим рождением обязано человеческой лени.
Содержание
- Преимущества автомобиля с гидроусилителем
- Устройство, схема ГУР
- Работа гидроусилителя
- Уход за гидроусилителем
Преимущества автомобиля с гидроусилителем
Как уже было отмечено, гидроусилитель рулевого управления изначально устанавливался на большегрузные автомобили, чтобы облегчить вращение руля. Кроме того усилитель делает меньшим передаточное отношение рулевого механизма, то есть водителю не приходится для совершения маневра делать баранкой 5-6 полных оборотов в сторону поворота и обратно. Это особенно выигрышно при выполнении парковки в стесненных условиях.
Кроме того усилитель делает меньшим передаточное отношение рулевого механизма, то есть водителю не приходится для совершения маневра делать баранкой 5-6 полных оборотов в сторону поворота и обратно. Это особенно выигрышно при выполнении парковки в стесненных условиях.
Гидравлика смягчает удары на руль от дорожных неровностей и помогает сохранить управляемость автомобилем при наезде на большой камень и даже при простреле передней шины.
Вскоре иностранные производители стали ставить их и на легковые машины, что сразу по достоинству было оценено потребителем.
К сожалению, отечественный автопром так и не наладил выпуск массовой модели легковых автомобилей с какой-нибудь схемой усиления руля. Правда, были собраны опытные единичные экземпляры (например, ГАЗ 13/14 «Чайка» с АКПП и гидроусилителем руля), но они не были предназначены для простых смертных, и об их устройстве в народе ходили только легенды и догадки.
Устройство, схема ГУР
Гидроусилитель руля представляет собой такую систему, которая работает по определенной схеме и состоит из следующих элементов:
- бачок для рабочей жидкости;
- соединительные шланги;
- распределитель;
- гидроцилиндр;
- насос.


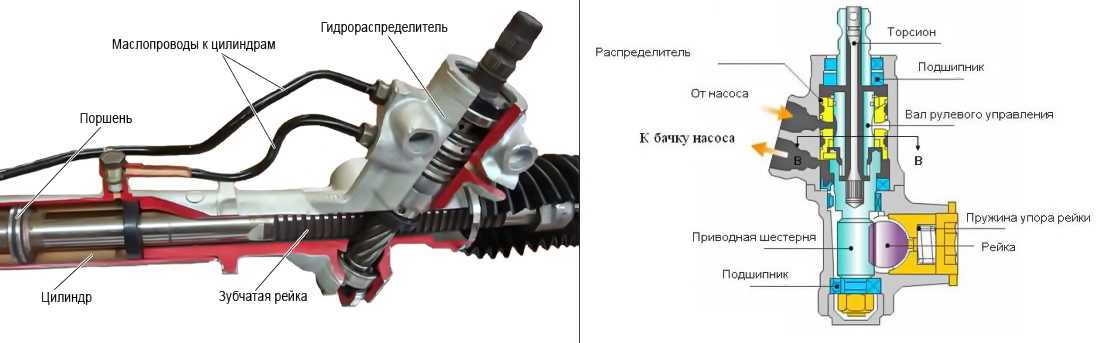
Насос предназначен поддерживать давление и обеспечивать циркуляцию рабочего масла в системе. Он закреплен на двигателе и приводится в действие ременной передачей от коленвала. На большинстве гидроусилителей установлены пластинчатые насосы.
Распределитель – очень высокоточный механизм. Он призван направлять потоки рабочей жидкости в необходимую полость гидроцилиндра, а после обратно в бачок. Устанавливается на элементах рулевого привода либо на валу рулевого механизма. Различают распределители осевые – если подвижный элемент (золотник) перемещается поступательно, и роторные – если подвижный элемент вращается.
Гидроцилиндр под давлением жидкости и посредством поршня и штока поворачивает колеса. Встраивается в рулевой механизм или может быть расположен между элементами рулевого привода и кузовом.
Бачок – резервуар для рабочей жидкости. В нём обязательно установлен фильтрующий элемент и находится щуп для контроля за уровнем масла. Это масло, кроме передачи усилий от насоса к гидроцилиндру, призвано смазывать все пары возникающего трения.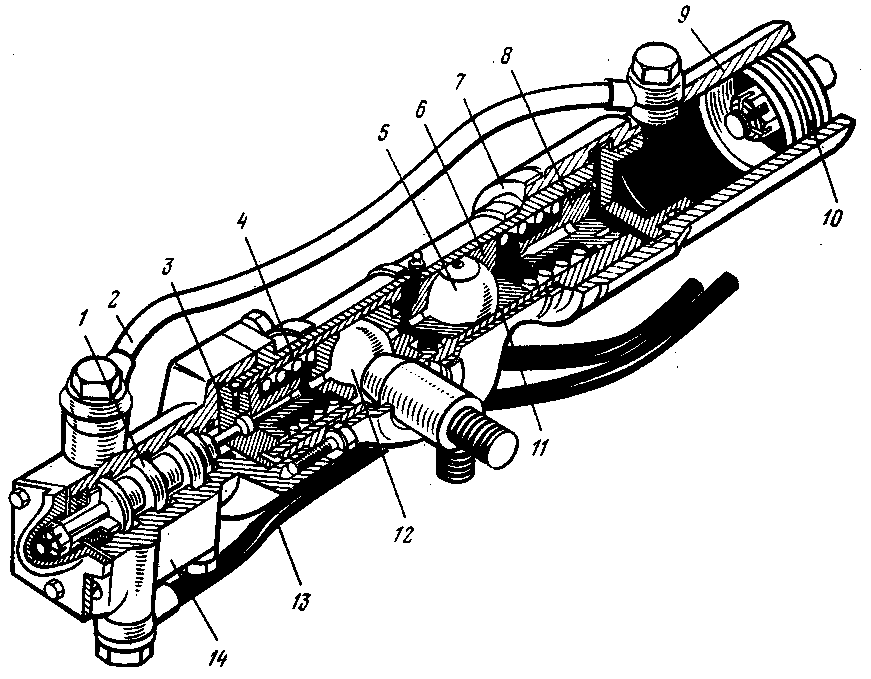
Соединительные шланги (высокого давления) обеспечивают циркуляцию масла между насосом, распределителем и гидроцилиндром. Из бачка в насос и из распределителя обратно в бачок гидроусилителя жидкость поступает по шлангам низкого давления.
Работа гидроусилителя
Принцип работы гидроусилителя руля, как с осевым, так и с роторным распределителем, основан на перемещении золотника при перекладке рулевого колеса. Перемещаясь, он перекрывает одну из сливных магистралей, и масло под давлением поступает в ту или иную (правую или левую) рабочие полости гидроцилиндра. Рабочая жидкость давит на поршень со штоком, и те в свою очередь поворачивают колеса. Колеса поворачивают за собой, в сторону движения золотника, корпус распределителя.
Как только перестает вращаться рулевое колесо, схема меняется. Останавливается золотник, и восстанавливается нейтральное положение корпуса распределителя. Жидкость без препятствий перетекает из нагнетательной магистрали в сливную.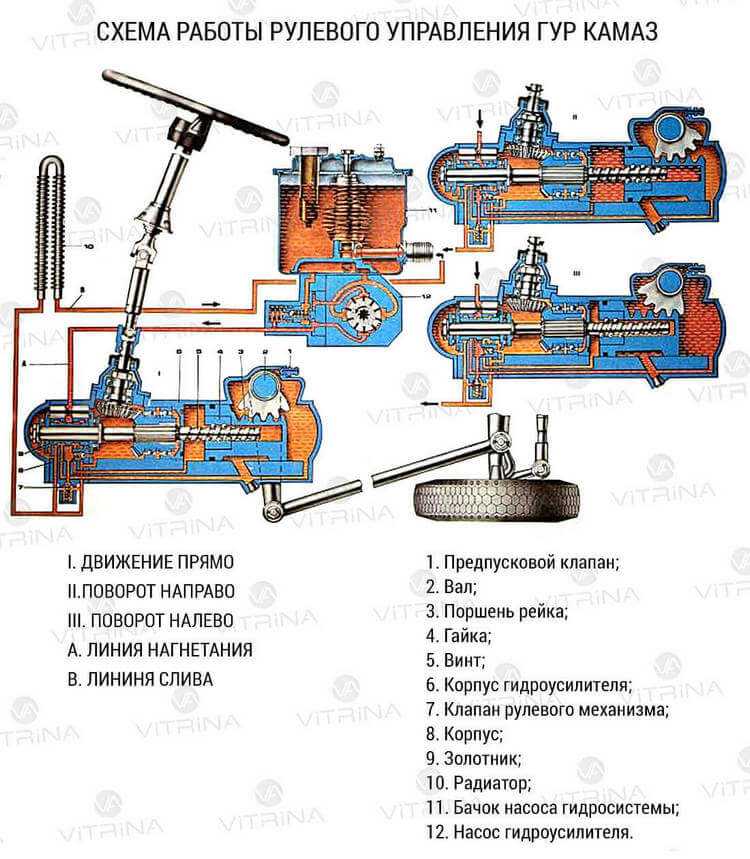 Усилитель рулевого управления с помощью насоса просто прокачивает масло по системе. Колеса стоят прямо.
Усилитель рулевого управления с помощью насоса просто прокачивает масло по системе. Колеса стоят прямо.
Необходимо отметить, что в случае выхода из строя гидронасоса, управление автомобилем не теряется.
Уход за гидроусилителем
Для надежной и бесперебойной работы устройство гидроусилителя руля требует постоянного контроля и ухода. Вот некоторые рекомендации.
- периодически проверяйте уровень масла в бачке;
- следите за герметичностью системы, своевременно устраняйте различные утечки;
- проверяйте натяжение приводного ремня и при необходимости регулируйте;
- раз в 1-2 года меняйте фильтрующий элемент в бачке. Следует своевременно производить замену масла, следите за изменением его цвета;
- помните, что на автомобиле с гидроусилителем нельзя удерживать руль в крайнем поворотном положении более 5-6 секунд. Это может привести к перегреву масла;
- не допускайте долгой эксплуатации автомобиля с неработающим гидронасосом. Это приведет к быстрому износу и выходу из строя распределителя и деталей рулевого механизма.

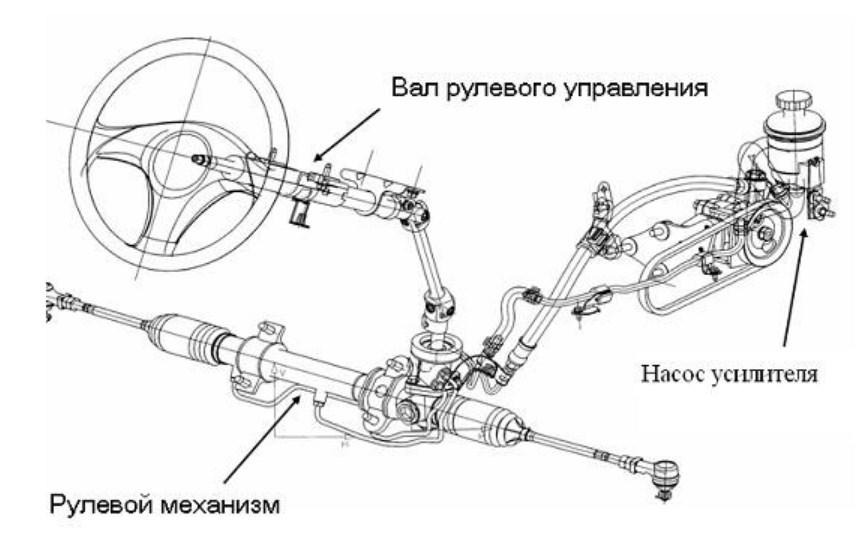
устройство и принцип работы, схема рулевого управления с ГУРом
Гидроусилитель руля (аббревиатура ГУР) — знакома большинству автолюбителей. Относится она к основной части рулевого механизма. Раньше управление машиной, было очень утомительным занятием, так как приходилось при резкой смене траектории напрягаться для поворота рулевого колеса, особенно это было проблематично на грузовых машинах. Конструкторы, которые всегда совершенствуют детали для удобства, комфорта и безопасности, обратили на это внимание, поэтому рулевое колесо стало не исключением. Чтобы свисти усилие к минимуму, была придумана система гидроусилителя руля.
Основным его предназначением, как и было, задумано выступает, комфортное управление машиной в момент движения, но есть и другие не менее важные заслуги, такие как:
- сохранение «обратной связи»;
- обеспечение устойчивости на дороге;
- повышение безопасности. То есть происходит контроль над ТС после повреждения передней шины и возможность увильнуть от столкновения;
- позволяет «чувствовать» дорожное полотно и создает кинематическое следящее действие;
- уменьшает передаточное отношение рулевой системы, что повышает маневренность;
- продлевает время службы деталей рулевого узла.
Гидроусилитель руля
По конструкции ГУР компактны и могут поглощать удары, с вибрацией отходящие от дорожного полотна на рулевое колесо. Во время использовании они совершенно бесшумны. С их появлением езда стала безаварийной, даже число парковочных мест снизилось вдвое. Имея в авто гидроусилитель сложные повороты и многоразовые маневры стали даваться на ура. Однако многие не знают принципа работы установленного штатного гидроусилителя, а когда транспортное средство уводит в сторону они пытаются разрешить ситуацию на «сход-развале» делая это неверно. Естественно, бывалые развальщики легко могут выставить углы установки колес для правильного «сопротивления» увода машины вбок, если гидроусилитель неисправен.
Чтобы устройство не подводило и надежно выполняло все предписанные задачи нужно своевременно посещать сервисные центры для диагностики.
Содержание
- 1 Шаг в историю
- 2 Разновидности гидроусилителей
- 3 Гидроусилитель руля: устройство и принцип работы
- 4 Устройство насоса гидроусилителя руля
- 5 Техническое обслуживание гидроусилителя
- 6 Недостатки ГУР
Шаг в историю
Так как первые машины по конструкции были не увесистыми и с узкими колесами, то для поворота руля не требовалось особых усилий.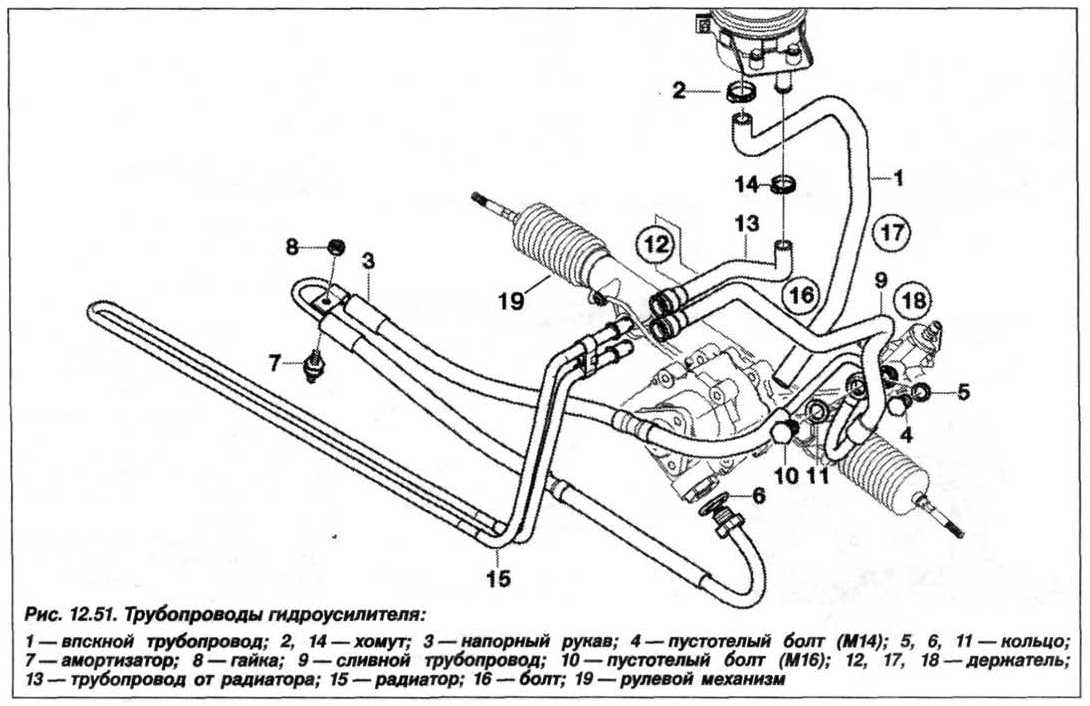 Но с появлением первых грузовых автомобилей вращать колеса многотонного грузовика, оказалось занятием достаточно трудоемким, а то и вовсе не посильным. Тут-то и потребовалось уменьшить диаметр «баранки» и изменить устройство рулевой рейки. Изобрел и запатентовал гидроусилитель впервые Фредерик Ланчестер. Сначала, благо автомеханики распространилось только на карьерные самосвалы, пожарные и грузовые машины. Предвестники пневмоусилители — были несложными и подпитывались от компрессора уже существующих пневматических тормозов.
Но с появлением первых грузовых автомобилей вращать колеса многотонного грузовика, оказалось занятием достаточно трудоемким, а то и вовсе не посильным. Тут-то и потребовалось уменьшить диаметр «баранки» и изменить устройство рулевой рейки. Изобрел и запатентовал гидроусилитель впервые Фредерик Ланчестер. Сначала, благо автомеханики распространилось только на карьерные самосвалы, пожарные и грузовые машины. Предвестники пневмоусилители — были несложными и подпитывались от компрессора уже существующих пневматических тормозов.
Только в 20-х годах XX-го века компания Rolls-Royse оснастила гидроусилителем машину-визитку Phantom. Понятное дело, гидравлические усилители были сложнее, чем уже существующие пневматические. Но попытка не увенчалась успехом, и эксперимент был отложен на несколько лет. Дальше уже во время Второй мировой войны англичане вновь ввели в работу ГУР, установив его на большие бронированные автомобили. И уже спустя пять лет технология плотно закрепилась в европейском и американском автопроме. С тех пор устройство не претерпевало принципиальных изменений. Сегодня разнообразие системы ГУРа впечатляет, помимо него существуют еще две удивительных технологии облегчающие эксплуатацию транспортных средств – Электроусилитель и Электрогидроусилитель.
С тех пор устройство не претерпевало принципиальных изменений. Сегодня разнообразие системы ГУРа впечатляет, помимо него существуют еще две удивительных технологии облегчающие эксплуатацию транспортных средств – Электроусилитель и Электрогидроусилитель.
Разновидности гидроусилителей
Утверждать, что ГУР в стандартном исполнении крайне необходим нельзя. Он полезен только в определенных моментах. Конечно, он позволяет с успехом маневрировать в городских условия, но вот на открытой трасе при высокой скорости пользы от него вовсе нет. С возрастанием скорости перестаёшь «чувствовать» дорогу, что популярно особенно в зимний период.
Чтобы, как-то перекрыть изъян, было предпринято установить рулевую рейку с переменным придаточным отношением. Однако попытка была безуспешной спасла ситуацию электроника, которая выступила модификацией гидроусилителя. Она сочетает не только комфорт, но и информативность руля. Электрогидроусилитель руля (ЭГУР) служит по тому же принципу, отличия — прибавка электронного блока и исполнительного электроклапана.
Бачок гидроусилителя
Гидроусилитель руля: устройство и принцип работы
Чтобы понять, как устроена конструкция ГУРа, рассмотрим схему гидроусилителя рулевого управления, состоящую из таких частей, как:
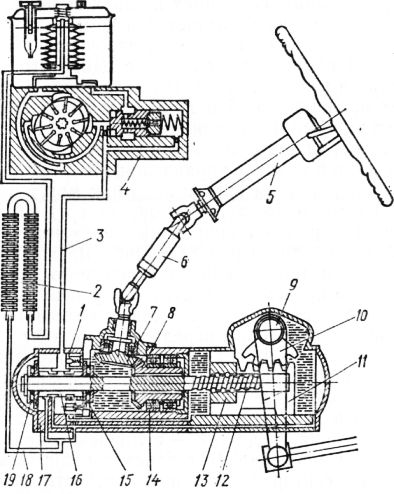
- Силовой гидроцилиндр двойного действия помещен в рулевую часть, где стоит межу деталями привода и кузова. Соединен он с золотниковым управляющим узлом и гидроцилиндром. Основная заслуга — преобразование давления жидкости в перемещение поршня и штока, помогающих двигать колеса в необходимое направление.
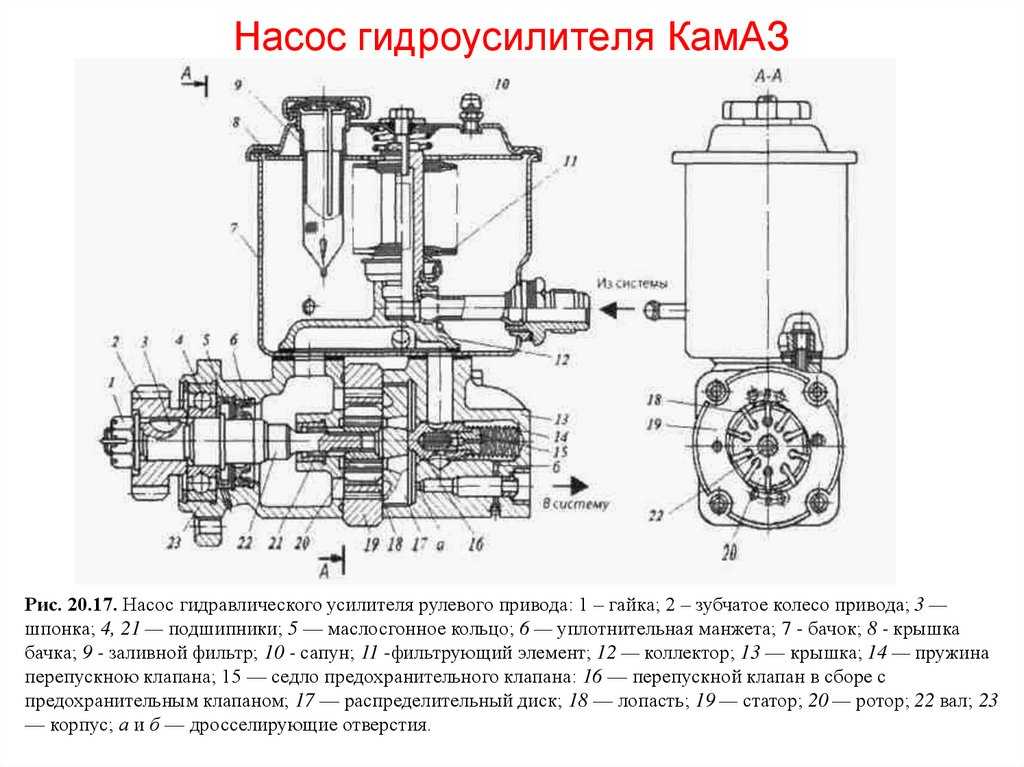
- Насос прикрепляется на двигателе, а его привод от коленчатого вала осуществляется ременной передачей от шкива коленчатого вала. Требуется для сформирования давления масла. Более распространены конструкции лопастого типа, потому что у них хорошее КПД.
- Рабочая жидкость содержится в бачке, там же имеется фильтр, крышка с щупом для замера уровня. Функция масла смазывать трущиеся детали и передавать усилие от насоса к гидроцилиндру.
- Бачок, наполненный гидравликой, чтобы содержать его в чистоте внутри есть фильтр.
- Регулятор давления или распределитель – это прецизионный (высокоустойчивый) и простой по схеме узел. Являет собой редукционный клапан. Располагается на деталях рулевого привода или на одном валу с рулевым элементом. Его задача распределять гидравлику в нужную полость гидроцилиндра или назад в бак. Требуется для контроля частоты вращения коленвала мотора, чтобы тот не повышал допустимую норму давления гидравлической жидкости. Золотниковый распределитель – сложная деталь, состоящая из торсиона и золотникового клапана. Когда находящийся внутри золотник крутится, распределитель называют роторным, а если поступательно перемещается – осевым.
- Соединительные шланги высокого и низкого давления сводят между собой гидроцилиндр, насос и распределитель. Также по ним циркулирует гидравлическая жидкость из бака в насос и обратно, возвращаясь от распределителя. Там, где требуется создать взаимную подвижность узлов, применяют гибкие шланги.
Принцип работы у ГУР и ЭГУР схож

 Там, где требуется создать взаимную подвижность узлов, применяют гибкие шланги.
Там, где требуется создать взаимную подвижность узлов, применяют гибкие шланги.Принцип работы гидроусилителя руля как с осевым, так и с роторным распределителем, основан на перемещении золотника при перекладке рулевого колеса. Сначала насос формирует давление в узле рулевого управления. Если «баранку» крутят в одну из сторон, начинает двигаться золотник и закрывает одну из сливных магистралей, а рабочая жидкость под давлением идет в нужные полости гидроцилиндра. Гидравлика со штоком давит на поршень, а он двигает колеса.
Когда колеса поворачиваются, они направляют корпус распределителя в сторону движения золотника. А когда золотник принимает обездвиженное состояние начинают восстанавливать свое обычное положение корпуса распределителя. Из нагнетательной магистрали масло легко проходит в сливную. Далее, усилитель просто качает рабочую жидкость при помощи насоса по системе. В то же время колеса направлены прямо. Когда руль заканчивает крутиться, вся схема меняется и останавливается.
Если даже гидронасос сломался (к примеру, оборвался ремень привода) — это не влияет на управление транспортным средством. Потому что от рулевой системы усилие будет идти на корпус распределителя, а после на колеса с золотником. Через предпусковой клапан, гидравлика станет двигаться из одной полости в другую и не создавать препятствия, чем позволит поворачивать руль, только с напрягом. Схема рулевого управления с гидроусилителем наглядно демонстрирует всю суть системы.
Устройство насоса гидроусилителя руля
Во время поворота рулевого колеса в другую сторону распределитель подает масло в противоположные части гидроцилиндра, соответственно рулевая рейка идет в другую сторону и поворачивает колеса в нужную сторону. Что касается водителя, то он прилагает минимум усилия на поворот руля. Когда автомобиль находится без движения руль поворачивать также просто для этого необходимо чтобы был запущен мотор.
Если транспортное средство наезжает на препятствие, сила отталкивания пытается повернуть колеса. Но вместо этого они относительно золотника двигают корпус распределителя и перекрывают сливную магистраль. После чего гидравлическая жидкость поступает в полость цилиндра, и поршень посылает усилия на колеса, идущие в обратном направлении. Быстрая реакция приводит к тому, что колеса блокируются и не могут поворачивать. Из-за того, что ход золотника малый (где-то 1 мм), транспортное средство практически не меняет направление движения. ГУР ограждает руки водителя от столкновения со спицами руля, когда он во что-то врезается. Маленькие толчки все-таки ощущаются – это происходит из-за того, что над реактивными шайбами, повышается давление.
Но вместо этого они относительно золотника двигают корпус распределителя и перекрывают сливную магистраль. После чего гидравлическая жидкость поступает в полость цилиндра, и поршень посылает усилия на колеса, идущие в обратном направлении. Быстрая реакция приводит к тому, что колеса блокируются и не могут поворачивать. Из-за того, что ход золотника малый (где-то 1 мм), транспортное средство практически не меняет направление движения. ГУР ограждает руки водителя от столкновения со спицами руля, когда он во что-то врезается. Маленькие толчки все-таки ощущаются – это происходит из-за того, что над реактивными шайбами, повышается давление.
«Чувство дороги» — это обратная связь от управляемых колес через усилитель к рулю. Сообщает водителю, в каких условиях происходит поворот. Чувствуя силовое следящее действие управлять машиной можно при любой погоде. Поэтому в составе конструкции крепят реактивные шайбы, плунжеры или камеры. Одна из шайб при высоком давлении, пытается поместить золотник в исходную точку, от этого рулевое колесо работает «туже».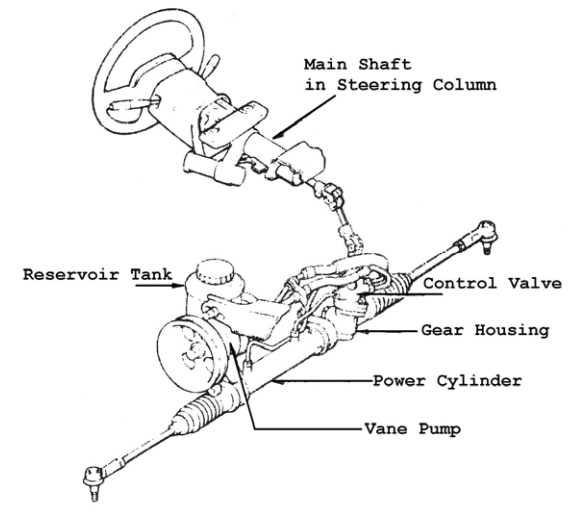
Расположение ГУРа
Устройство насоса гидроусилителя руля
Узел насоса лопастного типа делится на виды:
- Лопастный.
- Шестеренный.
Механизм насоса состоит из корпуса, ротора и уплотнительного кольца. Насос имеет клиноременный привод от шкива коленчатого вала. Шкив матируется в конце наружного вала, находящийся на шариковом и игольчатом подшипнике. Ротор располагается на шлицах вала, в его пазы свободно установлены лопасти. К корпусу насоса приделан распределительным диском и крышкой статор.
Внутренняя поверхность его корпуса имеет сложную форму. Лопасти устанавливаются в ротор, где параллельно его продольной оси предусмотрено несколько прорезей. Эти лопасти под давлением центробежной силы немного выходят из пазов и соприкасаясь, с внутренней поверхностью корпуса, создают замкнутые камеры.
Внутренняя поверхность корпуса устроена таким образом, что когда объём от вращения ротора снижается между ними сжимается масло.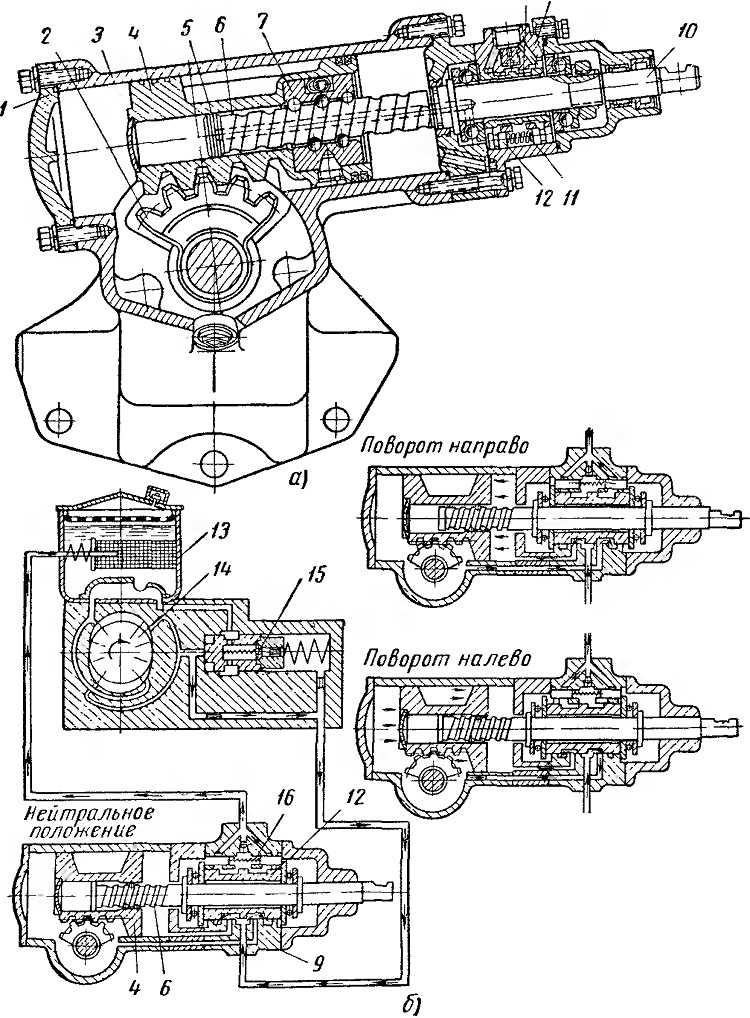 Если появляется отверстие, то гидравлическая жидкость стремительно выходит из лопастей. Процесс всасывания жидкости проходит наоборот. Сам по себе насос должен быть высокопроизводительным, чтобы обеспечивать повороты вала максимально быстро.
Если появляется отверстие, то гидравлическая жидкость стремительно выходит из лопастей. Процесс всасывания жидкости проходит наоборот. Сам по себе насос должен быть высокопроизводительным, чтобы обеспечивать повороты вала максимально быстро.
Запускается передачами от двигателя:
- Шестеренчатой.
- Ременной.
Рулевое управление с гидроусилителем, совмещенным с рулевым механизмом
Техническое обслуживание гидроусилителя
- Очень часто приводящий ремень становится причиной поломки гидроусилителя, поэтому нужно следить за уровнем его натяжения.
- Необходимо смотреть за уровнем масла в бачке, если его показатель ниже нормы, нужно долить. Нехватка масла приведет к тому, что насос выйдет из строя. Какая подходит гидравлическая жидкость больше всего нужно узнать из инструкции или в специализированном магазине.
- Один раз в год меняйте фильтрующий элемент в бачке.
- Сливать масло легко, нужно снять шлангу и при открытой крышке бачка жидкость выльется от действия атмосферного давления. Залив свежее масло необходимо при открытой крышке пару раз прокрутить руль в крайние положения, это нужно, чтобы лишний воздух покинул бачок.
- Учтите, что на ТС с гидроусилителем не стоит держать руль в крайнем поворотном положении свыше 5–6 секунд – перегреется гидравлическая жидкость.
- Кроме того, следует время от времени осуществлять замену масла, это объясняется загрязнениями, которые влияют на его свойства. При потере основных свойств жидкость способна повредить сальники рулевой рейки, а это приведет к сбою ГУРа.
- Регулярно проводите визуальный осмотр системы, так как может быть нарушена герметичность системы. Потеки гидравлики говорят, что требуется ремонт.
- Если гидронасос не работает, использовать машину долгое время нельзя – это приведет к износу распределителя и поломке элементов рулевого механизма.
Залив присадки в ГУР
 Залив свежее масло необходимо при открытой крышке пару раз прокрутить руль в крайние положения, это нужно, чтобы лишний воздух покинул бачок.
Залив свежее масло необходимо при открытой крышке пару раз прокрутить руль в крайние положения, это нужно, чтобы лишний воздух покинул бачок.Недостатки ГУР
- Нужно просматривать систему каждый день на наличие дефектов.
- Нанос работает от мотора, тем самым забирая у него часть мощности.
- Нет функций регулировки положений работы для разных условий.
Как видно минусов не так уж и много.
Интересное по теме:
загрузка…
Устройство гидроусилителя руля. | Диагностика двигателя
Управляемость автомобиля напрямую зависит от конструкции и состояния
узлов рулевого управления. Практически все современные автомобили
оснащаются гидроусилителем руля. Но не все автолюбители представляют
принцип работы гидроусилителя руля. В связи с этим, когда машину
начинает уводить в сторону, основная масса водителей пытается решить
проблемы неправильной работы рулевого управления на «сход-развале».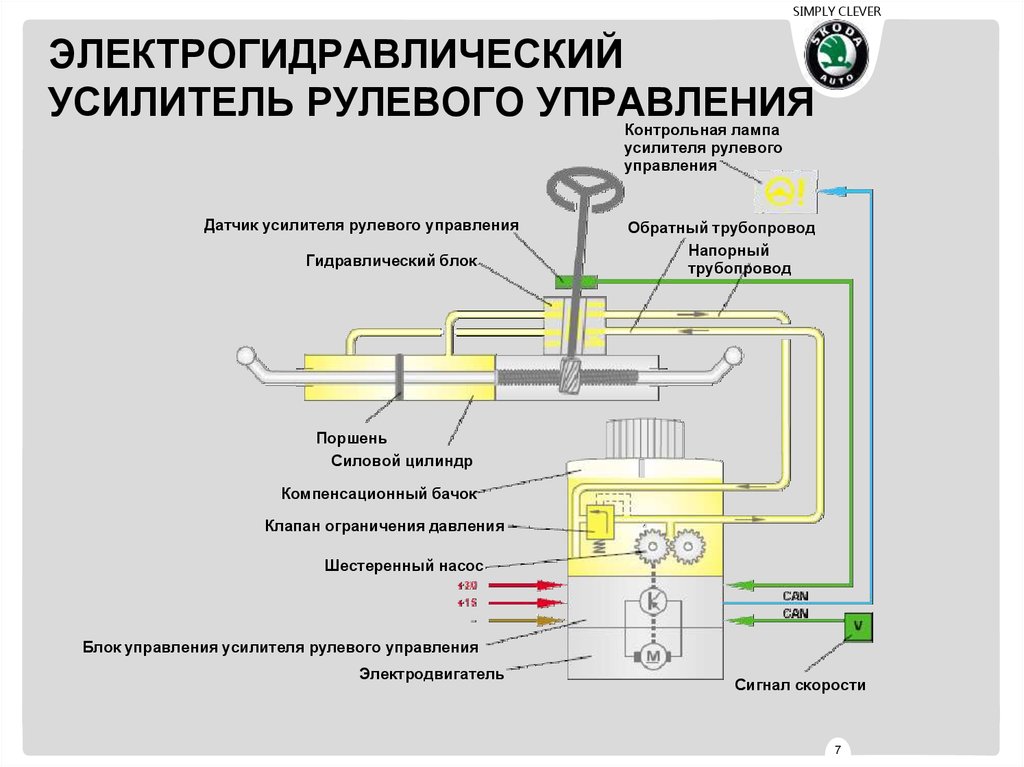 Конечно опытный развальщик может выставить углы установки колес таким
образом, что бы они «сопротивлялись» уводу автомобиля в сторону из-за
неправильной работы гидроусилителя. Например если неправильно работает
золотниковый механизм, то давление в силовом цилиндре при повороте
вправо и влево будет разным, а значит и усилие на руле будет разным.
Другой пример, при отсутствии усилия на руле (прямолинейное движение
автомобиля) рабочая жидкость все равно попадает в цилиндр под давлением
по одной магистрали высокого давления, при этом руль, а самое главное и
колеса, будет поворачиваться в сторону, при этом машину начинает тянуть.
Бывали случаи, когда на стенде сход-развала (на пятаках) на заведенной
машине, при отпущеном руле, колеса поворачивались сами до упора.
Конечно опытный развальщик может выставить углы установки колес таким
образом, что бы они «сопротивлялись» уводу автомобиля в сторону из-за
неправильной работы гидроусилителя. Например если неправильно работает
золотниковый механизм, то давление в силовом цилиндре при повороте
вправо и влево будет разным, а значит и усилие на руле будет разным.
Другой пример, при отсутствии усилия на руле (прямолинейное движение
автомобиля) рабочая жидкость все равно попадает в цилиндр под давлением
по одной магистрали высокого давления, при этом руль, а самое главное и
колеса, будет поворачиваться в сторону, при этом машину начинает тянуть.
Бывали случаи, когда на стенде сход-развала (на пятаках) на заведенной
машине, при отпущеном руле, колеса поворачивались сами до упора.
В золотниковом механизме (роторный управляющий клапан) совмещены
маслопровод подачи и стока. Гидравлическая жидкость перетекает из
трубопровода высокого давления в масляный резервуар, не выполняя никакой
работы
.
Конструкция и принцип функционирования элементов гидроусилителя рулевого управления — схема работы
Принцип действия реечного механизма с гидроусилителем. В корпусе рейки — торсионный стержень, связанный с рулевым валом. При повороте рулевого вала (колеса), стержень, поворачиваясь, перемещает золотник. Золотник приоткрывает отверстия каналов, идущих к силовому цилиндру. Цилиндр передвегает рейку, снижая усилие на руле. При отсутствии усилия на руле, ротор возвращается в исходное положение, а жидкость перепускается обратно в бачок.
Функциональная схема системы гидросусилителя руля
1 — Силовой цилиндр | 11 — Резервуар гидравлической жидкости |
Работа гидроусилителя рулевого механизма
1 — Поршень | 4 — Силовой цилиндр |
Общая информация
Привод рулевого насоса осуществляется непосредственно от двигателя с
помощью ремня.
При прямолинейном движении автомобиля чувствительный к изменениям
давления клапан-переключатель насосной сборки остается открытым,
обеспечивая сброс гидравлической жидкости обратно в резервуар
системы гидроусилителя руля..
За счет клапана регулировки расхода давление гидравлической жидкости
поддерживается практически постоянным при любых оборотах двигателя.
Под регулируемым напором гидравлическая жидкость подается по шлангу
А к роторному управляющему клапану.
При поворачивании рулевого колеса соединенный с валом ведущей
шестерни роторный клапан открывает гидравлический контур в
направлении, соответствующем направлению поворота колес и
гидравлическая жидкость по трубке А или В подается в соответствующую
(А или В) рабочую камеру.
Поскольку рулевой вал через роторный управляющий клапан механически
соединяется с валом ведущей шестерни, потери управления не
происходит даже в случае отказа системы гидроусиления.
Конструкция и принцип функционирования рулевого механизма
Основу гидравлической части рулевого механизма составляют
объединенные в общую сборку роторный управляющий клапан и силовой
цилиндр реечной передачи. Шток рулевой рейки в используемой
конструкции играет роль поршня в силовом цилиндре, сквозь роторный
клапан проходит вал ведущей шестерни. Рабочие камеры цилиндра и
роторного клапана соединены между собой посредством двух
гидравлических трубок.
Шток рулевой рейки в используемой
конструкции играет роль поршня в силовом цилиндре, сквозь роторный
клапан проходит вал ведущей шестерни. Рабочие камеры цилиндра и
роторного клапана соединены между собой посредством двух
гидравлических трубок.
Конструкция роторного управляющего клапана (золотниковый механизм)
1 — Торсионный стержень | |
Схема функционирования
роторного клапана при отпущенном рулевом колесе
1 — Камера А | 6 — V4 |
Схема функционирования роторного клапана при вращении рулевого колеса вправо
1 — Камера А | 4 — V2 |
Схема подключения рулевого
насоса
1 — Рулевой насос | 2 — Бачок гидравлической жидкости |
Схема функционирования
рулевого насоса
1 — Бачок ГУР | 5 — Клапан управления расходом |
Схема функционирования
чувствительного к изменению давления клапана при отпущенном рулевом
колесе
1 — К бачку гидравлической жидкости | 3 — Подаваемая под напором от насоса жидкость
(выше) |
Схема функционирования
чувствительного к изменению давления клапана при вращении рулевого
колеса
1 — К бачку гидравлической жидкости | 3 — Подаваемая под напором от насоса жидкость
(выше) |
Принцип функционирования
редукционного клапана насоса гидроусилителя руля
1 — К бачку ГУР | 5 — Давление жидкости, пропускаемой через
клапан управления расходом (ниже критического) |
Управляющий клапан состоит из
вращающегося вместе с рулевым валом ротора, ведущей шестерней, введенной
в зацепление с ротором посредством торсионного стержня и вращающейся
вместе с шестерней муфты. Конструкция клапана представлена на рисунке. В
роторе и муфте клапанной сборки предусмотрены канавки С и D, образующие
проходные каналы с V1 по V4 для потока гидравлической жидкости.
Конструкция клапана представлена на рисунке. В
роторе и муфте клапанной сборки предусмотрены канавки С и D, образующие
проходные каналы с V1 по V4 для потока гидравлической жидкости.
Нарушение исправности
функционирования системы гидроусиления (например, в результате обрыва
ремня) приводит к отказу повышения гидравлического давления, в
результате чего прикладываемый к рулевому колесу крутящий момент
начинает механически передаваться от ротора управляющего клапана
непосредственно на ведущую шестерню рулевого механизма. Но при этом
усилие не руле значительно увеличивается.
ремень ГУР, бачок, трубка, радиатор, сальник и другие детали
30.04.20221 914 1 1 ЭБУ
Автор:Иван Баранов
На сегодняшний день существует несколько видов систем усилителя руля, использующихся для обеспечения более простого управления рулем автомобиля. Одним из таких является гидроусилитель. Что представляет собой эта система, в чем заключается ее принцип работы, какие функции выполняет ремень ГУР? Ответы на эти и множество других вопросов вы сможете найти ниже.
Содержание
- 1 Экскурс в историю
- 2 Разновидности гидроусилителей
- 3 Конструктивные особенности
- 3.1 Фотогалерея «Основные элементы системы гидроусилителя»
- 4 Техническое обслуживание гидроусилителя
- 5 Недостатки ГУР
- 6 Видео «Как отремонтировать насос ГУР в гаражных условиях?»
[ Раскрыть]
[ Скрыть]
Экскурс в историю
Первые ГУРы стали ставиться на автомобили производителя Роллс-Ройс еще в 20-х годах прошлого века. Предвестником гидроусилителей были пневматические системы, которые по своей конструкции были более простыми. На тот момент гидроусилители не получили широкой популярности, в результате чего эксперимент с их использованием был отложен на какое-то время.
Повторно ГУРы были введены в эксплуатацию в период Второй Мировой войны, тогда они устанавливались на броневики. Через несколько лет эта технология стала повсеместно применяться в автомобилях европейского и американского производства. В принципе, с тех времен устройство и схема гидроусилетеля не претерпела серьезных изменений.
В принципе, с тех времен устройство и схема гидроусилетеля не претерпела серьезных изменений.
Разновидности гидроусилителей
Любой специалист скажет, что применение гидроусилителя более актуально при эксплуатации транспортного средства в условиях города — ее наличие дает возможность с большей легкостью совершать маневры. Однако при езде за городом, по трассе, большой пользы от ГУРа нет. Поскольку скорость движения по трассе обычно очень высокая, водитель авто с гидроусилителем попросту перестает чувствовать дорогу. А это очень опасно, особенно, если скорость движения действительно большая и особенно, в зимнее время года.
Для того, чтобы избавиться от этого недостатка, производители авто какое-то время использовали рулевые рейки с переменным придаточным отношением. Тем не менее, данный вариант не особо повлиял на недостатки ГУР, поэтому было принято решение о внедрении электроники в работе системы. Именно ее наличие стало модификацией, то есть определило подвид ГУР. Электрогидроусилитель выполняет те же функции и работает по аналогичному принципу, единственное различие заключается в добавлении управляющего модуля, а также исполнительного элемента — электрического клапана.
Электрогидроусилитель выполняет те же функции и работает по аналогичному принципу, единственное различие заключается в добавлении управляющего модуля, а также исполнительного элемента — электрического клапана.
Конструктивные особенности
Предлагаем ознакомиться с основными компонентами системы ГУР:
- Силовой гидравлический цилиндр двойного действия. Этот узел устанавливается в рулевую часть, в частности, между элементами привода и кузова. Он соединяется с золотником, а также гидроцилиндром. Назначение силового узла заключается в преобразовании давления расходного материала в перемещение штока и поршня. Последние используются для изменения положения угля колес.
- Бачок руля с подключенными трубками. В нем содержится гидравлическое масло. Для того, чтобы жидкость внутри резервуара была чистой, используется фильтрующий элемент.
- Радиатор ГУРа.
- Насосное устройство с сальниками и подшипниками, фиксируется на моторе. Привод насоса от коленвала производится посредством ременной передачи от шкива. Используется для образования давления рабочей жидкости.
- Смазочный материал, который находится в расширительном бачке. Назначение масла заключается в смазывании трущихся компонентов и дальнейшей передаче усилия от насоса на цилиндр.
- Регуляторное устройство или редукционный клапан. Этот элемент находится на деталях рулевой рейки либо привода. Предназначение клапана заключается в распределении гидравлической жидкости в соответствующую полость гидравлического цилиндра либо обратно в расширительный бачок. Также позволяет контролировать частоту вращения коленчатого вала двигателя, в противном случае последний может превысить допустимую норму давления масла. Конструктивно это достаточно сложная деталь, которая состоит из торсиона, а также золотникового клапана.
- Соединительные патрубки низкого и высокого давления, подключаются между собой цилиндр, насосное устройство, а также распределительный узел. По этим патрубкам осуществляется циркуляция смазывающего материала из резервуара в насос и обратно.
 Используется для образования давления рабочей жидкости.
Используется для образования давления рабочей жидкости.
Фотогалерея «Основные элементы системы гидроусилителя»
Что касается принципа действия, то он основывается на перемещении золотника при вращении рулем. Когда водитель вращает рулевое колесо, насосное устройство образовывает давление в системе, после чего происходит перемещение золотника. Последний закрывает один из сливных шлангов, а смазка начинает поступать в соответствующие полости цилиндра. Посредством гидравлики, которая со штоком давит на поршень, происходит изменение угла положения колес.
Когда колеса автомобиля поворачиваются в определенную сторону, они позволяют направить корпус распределительного узла в сторону перемещения золотника. В тот момент, когда золотник перестает двигаться, корпуса распределительного узла переводятся в исходное положение. Из нагнетательных шлангов смазывающая жидкость поступает в сливную магистраль, после чего усилитель осуществляет перекачку расходного материала с помощью насоса. В это время колеса автомобиля направлены прямо, а когда они поворачиваются, вся схема изменяется и останавливается.
В том случае, если насос выйдет из строя либо оборвется приводной ремешок, это не отразится на функциональности системы и управлении авто. Все потом, что усилие от руля будет поступать на корпус распределительного узла, а затем — на колеса. Гидравлика начнет передвигаться из одной плоскости в другую через предпусковой клапан, при этом она не станет создавать препятствия, что позволит также поворачивать рулевое колесо. Единственная разница в том, что водителю придется приложить для этого небольшое усилие (автор видео — Саня Маевский).
Техническое обслуживание гидроусилителя
Что нужно знать о техническом обслуживании гидроусилителя:
- Приводной ремень автомобиля с кондиционером или без него является одной из причин, по которым ГУР выходит из строя. Автовладелец должен всегда следить за состоянием ремешка и его натяжением. На ремне не должно быть следов трещин и других дефектов.
- Также нужно следить за уровнем жидкости в резервуаре, поскольку если объем масла будет низким, смазку обязательно придется добавлять. При недостатке жидкости может произойти выход из строя насосного устройства.
- Надо следить за состоянием фильтра, расположенного в расширительном бачке. Хотя бы раз в год его следует менять.
- Если вы заметили, что в масле появился осадок, то расходный материал обязательно подлежит замене. Дело в том, что наличие любой грязи отражается на свойствах жидкости в целом. Когда смазка теряет свои характеристики, это может привести к повреждению сальников рейки, что в свою очередь может привести к сбою в работе гидроусилителя. Для слива необходимо отсоединить патрубок и открыть крышку резервуара, это позволит быстро слить жидкость из системы. После залива новой смазки нужно при открытой пробке бачка провернуть рулевое колесо в оба крайних положения. Это позволит избавиться от воздушной пробки.
- Необходимо учитывать, что в автомобилях с ГУР ни в коем случае нельзя удерживать рулевое колесо в крайних положениях на протяжении более 5 секунд. Это приводит к перегреву смазки в системе.
- Также каждый автовладелец должен время от времени производить визуальную диагностику системы, в частности, следить за тем, чтобы она была герметична. Если вы заметили на шлангах следы потеков, это свидетельствует о том, что система нуждается в ремонте. Все поврежденные патрубки подлежат замене.
- В том случае, если гидравлический насос вышел из строя, специалисты не рекомендуют длительно эксплуатировать транспортное средство. Это может стать причиной ускоренного износа распределительного узла и выходу из строя компонентов системы рулевого управления (автор видео — канал Ремонт Гидравлики).
 Это приводит к перегреву смазки в системе.
Это приводит к перегреву смазки в системе.Недостатки ГУР
Какие недостатки характерны для ГУРов:
- Каждый автовладелец должен время от времени проверять систему на предмет повреждений и различных дефектов.
- Насосное устройство функционирует от силового агрегата, в результате чего забирает у двигателя часть мощности.
- Системы гидроусилителя не предусматривают функцию регулировки положений работы при различных режимах эксплуатации авто.

Загрузка …
Видео «Как отремонтировать насос ГУР в гаражных условиях?»
Подробная пошаговая инструкция по ремонту насоса гидроусилителя руля, в частности, по замене сальника и подшипника, приведена в ролике ниже (автор видео — канал Своими руками).
Была ли эта статья полезна?
Спасибо за Ваше мнение!
Статья была полезнаПожалуйста, поделитесь информацией с друзьями
Да (100.00%)
Нет
Как работает гидроусилитель руля: устройство и схема работы
Содержание статьи
- Функционал и назначение
- Предъявляемые требования
- Особенности устройства
- Ось для легкового прицепа: виды, как сделать своими руками
- Как это работает
- Чем отличается климат контроль от кондиционера: схемы работы
- Прицеп Центр: где купить прицеп, официальный сайт
- Как часто менять жидкость
- Признаки неисправности помпы (водяного насоса): замена своими руками
- Плюс и минусы
- Фаркоп Паджеро Спорт: какой лучше выбрать, варианты конструкции
Всем привет! Сегодня говорим о том, как работает гидроусилитель руля на автомобилях.
В действительности сейчас сложно себе представить автомобиль, который не был бы оснащен усилителем руля. Кстати, тут мы уже сравнивали ГУР и ЭУР, потому к этому вопросу возвращаться не станет.
Наиболее распространенным остается именно гидравлический узел. Это устройство может применяться на большом количестве авто. Хотя на Ниве, на Опель, автомобилях Тойота, КАМАЗ, УАЗ, Газель и многих других. Что уж говорить, если ГУР ставится даже на ЗИЛ 130. Думаю, некоторым из вас хорошо знакомы такие усилители как ЮМЗ 6 или МТЗ 80.
Но сейчас немного не об этом.
В рамках нашего материала хочется узнать что-то новое и полезное, проверить ваши знания в области усилителей рулевого управления на авто. Возможно, в отдельном материале поговорим про все возможные неисправности ГУР. К примеру, почему руль плохо или туго вращается, клинит при повороте в одну сторону, работает рывками и пр.
Функционал и назначение
ГУР входит в состав рулевого стандартного управления транспортного средства.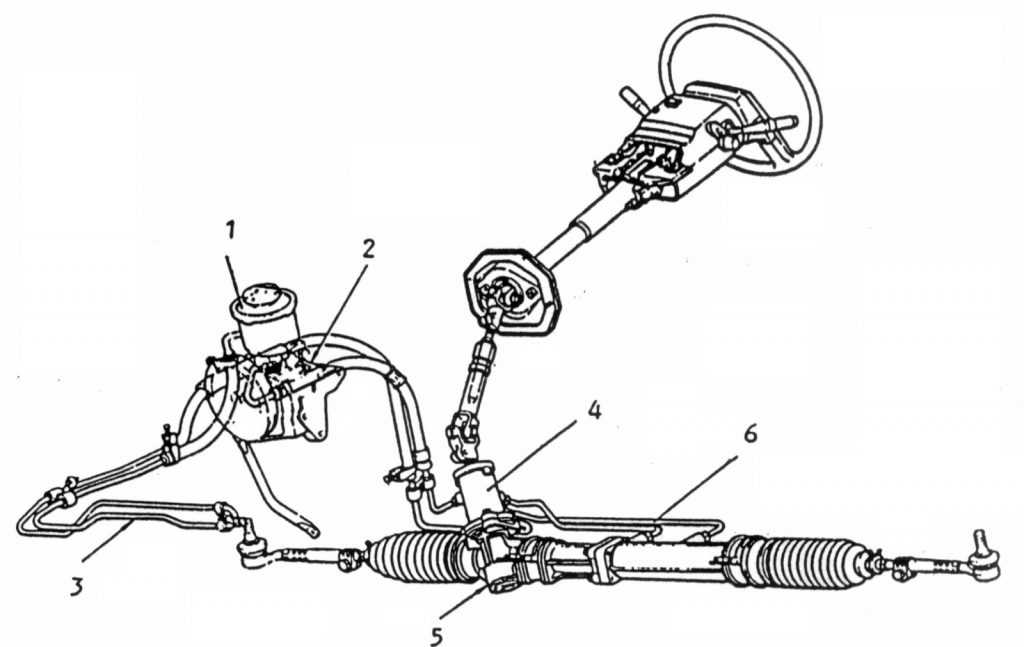 Он обеспечивает создание дополнительного, необходимого водителю, усилия при повороте автомобильного руля. Это реализуется за счет создаваемого гидравлического давления.
Он обеспечивает создание дополнительного, необходимого водителю, усилия при повороте автомобильного руля. Это реализуется за счет создаваемого гидравлического давления.
В случае с легковыми авто перед ГУР стоит основная задача. Это обеспечение комфорта во время управления. Рулить машиной с таким узлом намного проще, легче и удобнее. Для поворота или маневра совершать по несколько полных оборотов руля не приходится. А это наиболее сильно проявляется на узких участках дороги, на парковке и пр.
Также устройство ГУР позволяет сохранять управляемость и смягчать получаемые при наезде на неровности удары в рулевое колесо. Или просто в баранку.
Предъявляемые требования
Думаю, если вам этот материал окажется полезным, будет создана еще одна статья. В ней мы говорим о том, какие неполадки могут случаться с этим узлом.
К примеру, мы уже отдельно изучали причины скрипа руля при вращении. Еще отдельно рассматривался вопрос, когда руль не возвращается в исходное положение.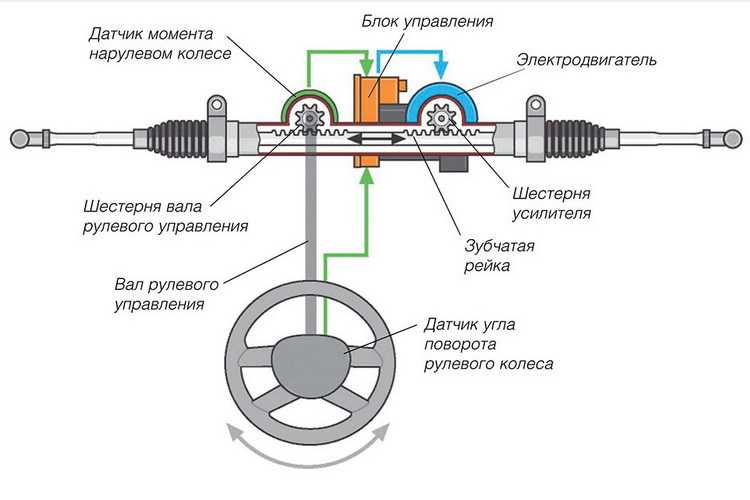 В этих случаях потенциальным виновником также часто выступает гидрач.
В этих случаях потенциальным виновником также часто выступает гидрач.
Чтобы ГУР обеспечивал надежную и качественную работу, он должен соответствовать определенным требованиям. Среди них:
- надежность и качество сборки;
- бесшумность;
- простота даже самостоятельного обслуживания;
- минимально возможные габариты;
- экологичность;
- автовозврат руля в исходное положение;
- плавность;
- пропорциональность усилий;
- сохранение работоспособности рулевого узла при поломке ГУР и пр.
Действительно важные требования. И если даже ГУР перестал им соответствовать, у водителя должна сохраняться возможность доехать до ближайшего СТО или своего гаража, к примеру. А не просто стоять и на холостых оборотах дожидаться эвакуатор.
Особенности устройства
Теперь поговорим непосредственно про устройство рассматриваемого узла.
Важным преимуществом системы является то, что ее можно поставить на рулевые механизмы различных типов. В случае с легковушками наиболее распространенными остаются реечные узлы.
В случае с легковушками наиболее распространенными остаются реечные узлы.
Если отталкиваться от такого механизма, тогда схема самого ГУР будет состоять из:
- бачка, внутри которого находится рабочая жидкость;
- насоса;
- золотникового распределителя;
- гидроцилиндра;
- соединительных шлангов.
Бачок представляет собой емкость, где находится рабочая жидкость. А именно специальное масло. Резервуар включает в себя фильтр и щуп, которым проверяется уровень.
Само масло отвечает за смазывание трущихся элементов, а также передает усилие от самого насоса непосредственно к цилиндру. То есть гидроцилиндру, как вы поняли.
Насос служит для поддержания правильного давления и обеспечения циркуляции жидкости. Причем этот узел ставится на блок цилиндров мотора, и работает от шкива коленвала. Сами насосы могут отличаться по своей конструкции. В основном это корпус из металла, внутри которого находятся вращающиеся на роторе лопасти.
Управление насосом может быть регулируемым, либо же нерегулируемым. В первом случае меняется производительная часть помпы. А во втором в игру вступает редукционный клапан.
В первом случае меняется производительная часть помпы. А во втором в игру вступает редукционный клапан.
Распределитель монтируют на рулевой вал, либо на компоненты привода рулевого узла. Именно он служит для правильного направления потоков рабочего масла в нужную полость гидроцилиндра, а также возвращают жидкость в бак.
Распределитель в основном состоит из торсиона, вала и не менее важного поворотного золотника. Узел бывает роторным и осевым.
Гидроцилиндр монтируется в рейку рулевого узла. Он включает в себя такие компоненты как поршень и шток. Шток меняет свое положение под воздействием масла.
С соединительными шлангами, думаю, все прекрасно понятно.
Как это работает
Чтобы правильно понять не только устройство, но и принцип работы, на котором основан ГУР рулевого управления, следует рассмотреть несколько наглядных ситуаций. Плюс еще видео поможет визуализировать многие моменты.
Представив несколько ситуаций, в каждой из них коротко объясним, что при этом происходит с усилителем.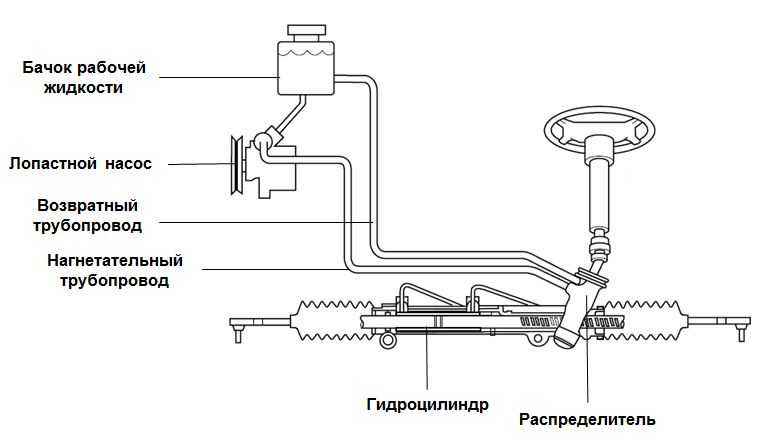
- Машина неподвижная, колеса выставлены прямо. Сейчас ГУР не работает и спецжидкость просто постепенно перекачивается по всей системе, от бачка до распределителя, и назад;
- Руль начинает вращаться. Крутящий момент от баранки идет на вал распределителя, а затем на торсион, начинающий закручиваться. Золотник при этом не вращается, что обусловлено силой трения. При перемещении относительно представленного золотника, рассматриваемый вал открывает специальный канал. Это позволяет жидкости начать поступать в определенную полость цилиндра. Полость напрямую зависит от того, в какую сторону происходит вращение. Жидкость начинает давить на поршень с штоком, что и позволяет сместить рулевую рейку и достаточно легко повернуть колеса влево или вправо;
- Вращение закончилось, но руль остается в повернутом положении. При перемещении рейка вращает золотник, и происходит его выравнивание в отношении вала. А именно вала рассмотренного распределителя. Теперь распределитель оказывается в нейтралке, и жидкость начинает просто продолжает циркулировать по системе. Никакая серьезная работа в этот момент не совершается;
- Руль оказывается в крайнем положении и удерживается. Это самая сильная нагрузка на ГУР. Связано с тем, что распределитель не способен вернуться в нейтраль, и жидкость активно циркулирует в самом насосе. От этого появляется шум. Как только руль будет отпущен, все нормализуется.
 Никакая серьезная работа в этот момент не совершается;
Никакая серьезная работа в этот момент не совершается;Важным преимуществом рассматриваемого узла является то, что рулевой механизм все равно сохранит свою работоспособность, даже если сам ГУР выйдет из строя.
Как часто менять жидкость
Довольно популярный вопрос. Нельзя сказать, что масло для ГУР дорогое. Нет, цена вполне адекватная, и во многом зависит от конкретного авто.
В теории использовать это масло можно весь срок. По факту лучше периодически проводить замену.
Периодичность замены напрямую связана с интенсивностью эксплуатации. К примеру, если за год вы проезжаете около 10-15 тысяч километров, сливать и заливать масло в усилитель можно с интервалом 2-3 года.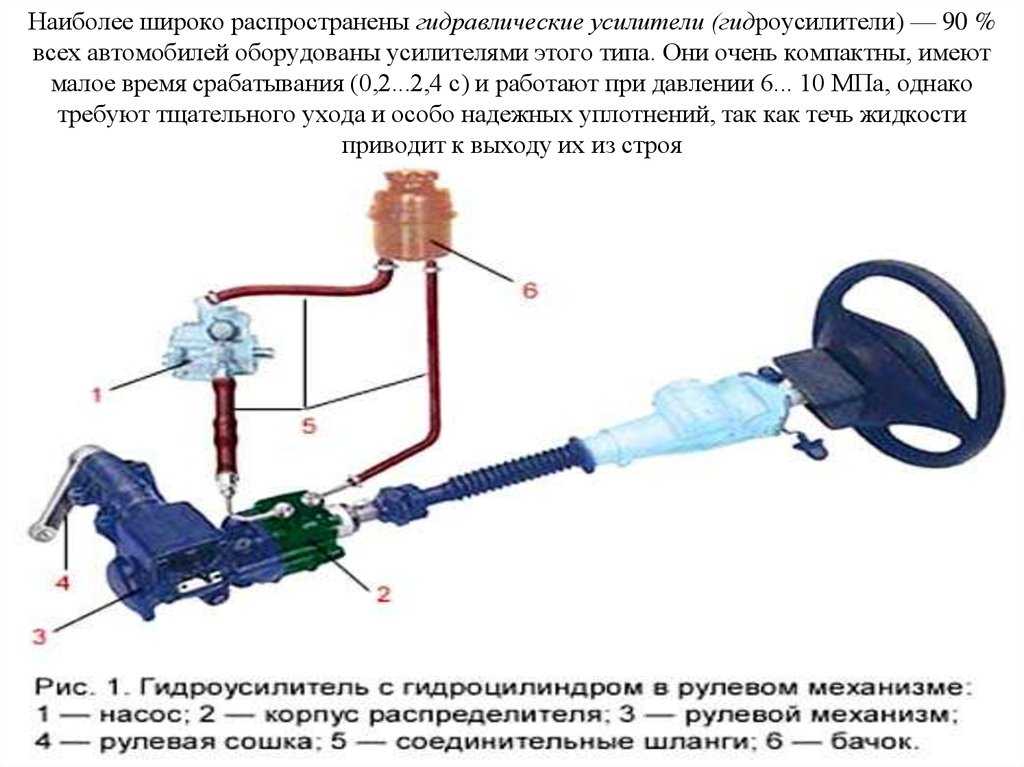 Если это 30-50 тысяч км, то интервал может составлять и 6-12 месяцев.
Если это 30-50 тысяч км, то интервал может составлять и 6-12 месяцев.
Если не менять состав, он будет постепенно терять свои свойства и уже не сможет справляться со своими обязанностями. В итоге начнется активный выход из строя всех компонентов.
А после замены нужно прокачать гидроусилитель, о чем мы с вами уже ранее говорили в предыдущих материалах. Там ничего сложного нет.
Плюс и минусы
Чтобы подвести итоги, стоит поговорить о сильных и слабых сторонах этого узла.
Начнем с преимуществ. Они такие:
- легкое управление;
- снижение усталости водителя;
- амортизация ударов неровной дороги;
- улучшенная маневренность;
- повышенная безопасность;
- более чувствительная управляемость.
Но есть и минусы. Для начала насос, который работает постоянно, будет забирать часть мощности у двигателя. Плюс это обслуживаемая система.
Насколько это весомые аргументы конкретно в вашем случае, решайте сами. И для наглядности советуем сравнить все же ГУР и ЭУР.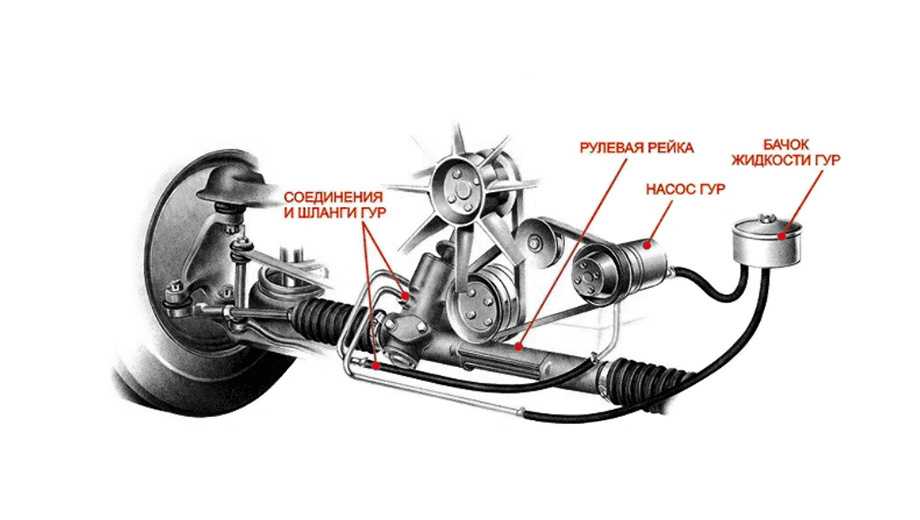 После чего сделать выводы.
После чего сделать выводы.
Watch this video on YouTube
На этом у нас все. Всем спасибо за внимание!
Подписывайтесь, оставляйте свои комментарии, задавайте актуальные вопросы!
Гидроусилитель руля: назначение и устройство
Для чего нужен ГУР? Большинство автолюбителей ответят: “Для того, чтобы легче крутить руль”. И будут правы, но отчасти. Кроме повышения комфорта, гидроусилитель позволяет уменьшить передаточное число рулевого управления. Что это дает? Чем больше передаточное число, тем меньшее усилие нужно прилагать для поворота колес. Но количество оборотов руля от упора до упора при этом будет равным 4-5. Уменьшая передаточное число, можно довести количество оборотов руля до 2-3. Управляемость, маневренность и острота реакций автомобиля улучшается, что особенно важно в аварийной ситуации, когда может не хватить времени для вращения руля с перехватами. Кроме того, у гидроусилителя есть еще несколько и преимуществ, и недостатков, о которых будет сказано ниже.
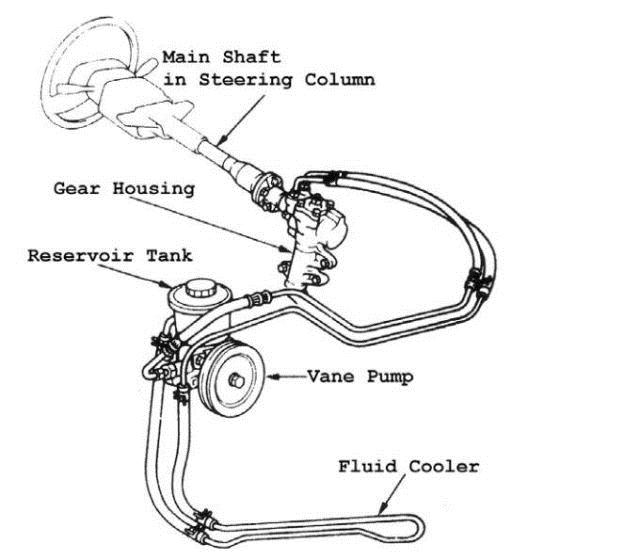
Гидроусилитель может устанавливаться на автомобили с рулевым управлением разных типов: червячным, винт-шариковая гайка. Мы расмотрим самый распространенный вариант – рейку. В состав системы гидроусиления входят:
- насос;
- распределитель;
- силовой цилиндр;
- бачок и соединительные шланги.
Насос гидроусилителя, как и любой другой насос, предназначен для создания и поддержания необходимого давление в системе и циркуляции рабочей жидкости (специального масла). Конструкция насоса может быть разной. Самые распространенные – лопастные, характеризующиеся высоким к.п.д. и износоустойчивостью. Насос крепится на двигателе и приводится в действие с помощью ремня от коленвала.
Распределитель, в зависимости от положения руля, направляет поток жидкости в соответствующую полость силового цилиндра или обратно в бачок. Он устанавливается на рулевом валу. Основные части распределителя – золотниковый клапан и торсион. Клапан состоит из двух цилиндрических частей с каналами для жидкости: внешней и внутренней. Торсион – это тонкий пружинистый металлический стержень, способный закручиваться под действием крутящего момента. Один конец торсиона соединен с рулевым валом, а второй – с шестерней, входящей в зацепление с рейкой. Внутренняя часть золотникового клапана соединяется с верхней частью торсиона, а внешняя – с его нижней частью.
Силовой цилиндр встроен в рейку. Он состоит из поршня и штока, перемещающего рейку под действием давления жидкости.
Рабочая жидкость передает усилие от насоса через распределитель к силовому цилиндру и смазывает все пары трения. Резервуаром для жидкости служит бачок. В нем может быть расположен фильтр, а в пробке — щуп для измерения уровня. Шланги высокого давления соединяют насос, распределитель и силовой цилиндр, а по шлангам низкого давления жидкость поступает в насос из бачка и возвращается в него из распределителя.
Содержание статьи
- 1 Принцип действия
- 2 Преимущества и недостатки
- 3 Электрогидравлический усилитель
Принцип действия
Как все это работает? Когда руль неподвижен (автомобиль стоит на месте, или движется по прямой), и система гидроусиления не задействована, в распределителе совмещены маслопроводы подачи и стока. Жидкость вхолостую перекачивается насосом через распределитель обратно в бачок. Когда водитель поворачивает руль, тем самым он закручивает торсион, а вместе с ним крутится и внутренняя часть золотникового клапана. Внешняя же часть пока остается неподвижной. Таким образом совмещаются каналы подачи жидкости в соответствующую полость силового цилиндра (в зависимости от того, в какую сторону повернут руль). Из другой полости силового цилиндра жидкость по открывшимся каналам сливается в бачок.Чем на больший угол повернут руль, тем сильнее закручивается торсион. Поэтому большим оказывается и размер перепускного отверстия, а, значит, и усилие, воздействующее на рейку. Рейка, перемещаясь, раскручивает через шестерню нижний конец торсиона, а вместе с ним и внутреннюю часть золотника. Обе части клапана возвращаются в исходное положение, и жидкость вновь перекачивается через распределитель в бачок.
В случае отказа системы гидроусиления потери управления не происходит, поскольку рулевой вал через торсион механически соединен с ведущей шестерней. Согласно нормам безопасности усилие на рулевом колесе легкового автомобиля не должно превышать 15 кг для полностью работоспособной и 30 кг — для неисправной системы рулевого управления. Быстродействие усилителя должно быть таким, чтобы при скорости вращения руля не менее полутора оборотов в секунду его не «закусывало».
Преимущества и недостатки
К перечисленным выше преимуществам ГУРа можно добавить смягчение ударов, передаваемых на руль от неровностей дороги и более четкое удержание автомобиля на выбранной траектории. Каким образом это происходит? Если, наехав на препятствие, колеса стремятся повернуться в сторону, это вызывает смещение рулевой рейки, ведущей шестерни и закручивание нижней части торсиона. Срабатывает золотниковый клапан, но “в обратную сторону”, так как усилие пришло не от руля, а от колес. Поэтому система будет не способствовать повороту колес, а противодействовать ему. То же самое происходит и при внезапном проколе шины: ГУР помогает автомобилю сохранять траекторию, а водителю – удержать руль в руках. Таким образом, усилитель повышает безопасность движения, а за счет повышения комфортности вождения снижает утомляемость водителя.
А теперь о недостатках. Во-первых, постоянно работающий насос отбирает часть мощности двигателя, даже когда ГУР не задействован. Причем производительность насоса должна быть такой величины, чтобы обеспечить легкий поворот колес на стоящем автомобиле – когда сопротивление максимально. Но ведь большую часть времени автомобиль движется, и усилий для поворота колес при этом нужно намного меньше! Вот и получается, что значительная часть отобранной у двигателя мощности пропадает впустую.
Во-вторых, производительность насоса зависит от оборотов двигателя – чем они выше, тем большее давление создает насос. А по идее все должно быть как раз наоборот – при малых скоростях движения необходимо максимальное усиление, а при высоких – небольшое. В простом гидроусилителе отсутствует возможность регулирования коэффициента усиления.
Из этого обстоятельства проистекает третий недостаток – противоречие между коэффициентом усиления и информативностью руля. Легкость и комфортность управления на малых скоростях имеет обратную сторону – “пустоту” руля на больших. Машина слишком “остро” реагирует на каждое движение руля, а отсутствие ощущения сопротивления (“обратной связи”) при повороте колес не дает возможности водителю правильно оценить их положение. Отчасти решить проблему помогают рейки с переменным передаточным отношением: в центре шаг зубьев небольшой, а к краям увеличивается. В этом случае при малых углах поворота машина не так остро реагирует на действия рулем, что очень важно на больших скоростях, зато на развороте крутить руль приходится меньше. Чем плох этот вариант? А тем, что передаточное отношение зависит от угла поворота руля, а не от скорости движения. Поэтому конструкторы стали искать другие пути.
Электрогидравлический усилитель
ЭГУР с электромагнитных клапаномЭГУР с электронасосомСхема работы ЭГУР с электронасосомНа помощь механике и гидравлике, как всегда, пришла электроника. В результате такого симбиоза появился электрогидравлический усилитель. Впервые его применили на автомобилях “Аudi” под названием “Servotronic”. Существует два типа ЭГУРа: с электромагнитным клапаном и с электронасосом. Управляет работой усилителя электронный блок на основании показаний датчиков скорости, поворота руля, оборотов коленвала. Набор датчиков может меняться в зависимости от модели автомобиля.
В первой конструкции в распределитель ГУРа дополнительно встраивается электромагнитный клапан и камера обратного действия с поршнем. При повороте колес на месте или при движении с малой скоростью клапан открыт, давление в системе максимально – руль крутить легко. При наборе скорости клапан, управляемый блоком, пропорционально закрывается. В результате давление в системе уменьшается, а усилие на руле увеличивается. Таким образом, получаем искомое чувство “обратной связи”.
Во второй, более совершенной конструкции, гидронасос заменен электронасосом, т.е. приводится не от коленвала, а отдельным электромотором. Управляет его работой опять же блок управления. На малых скоростях скорость вращения насоса максимальна, а на больших – ограничивается блоком управления. Поэтому чем выше скорость движения – тем “тяжелее” становится руль. Замена гидронасоса электронасосом позволяет снизить расход топлива до 0,2 л на 100 км.
Настраивая программу работы блока управления, можно адаптировать ЭГУР к различным моделям автомобилей. Более подробно о конструкции и принципе действия электрогидравлического усилителя можно прочитать здесь (формат PDF).
Нормальное развитие мозговых цепей
Скачать PDF
- Опубликовано:
- Грегори З. Тау 1 и
- Брэдли С. Петерсон 1
Нейропсихофармакология том 35 , страницы 147–168 (2010)Процитировать эту статью
27 тыс. обращений
684 Цитаты
46 Альтметрический
Сведения о показателях
Abstract
Охватывая функции от простейшей рефлекторной дуги до сложных когнитивных процессов, нейронные цепи выполняют разнообразные функциональные роли. В коре головного мозга функциональные области, такие как визуальная обработка, внимание, память и когнитивный контроль, зависят от развития отдельных, но взаимосвязанных наборов анатомически распределенных корковых и подкорковых областей. Организация развития этих цепей представляет собой удивительно сложный процесс, на который влияют генетическая предрасположенность, события окружающей среды и нейропластические реакции на эмпирические потребности, которые модулируют связи и коммуникации между нейронами в отдельных областях и цепях мозга, а также между нервными путями. Недавние достижения в нейровизуализации и вычислительной нейробиологии вместе с традиционными исследовательскими подходами, такими как гистологические исследования, клеточная и молекулярная биология, сыграли неоценимую роль в улучшении нашего понимания этих процессов развития у людей как в здоровом, так и в болезненном состоянии. Чтобы контекстуализировать истоки развития широкого спектра нейропсихиатрических заболеваний, в этом обзоре описывается развитие и созревание нейронных цепей от первого синапса через критические периоды уязвимости и возможностей до возникающей способности к когнитивной и поведенческой регуляции и, наконец, динамическое взаимодействие между ними. уровни схемной организации и эпохи развития.
ВВЕДЕНИЕ
Нейронные цепи, возможно, являются первичными надклеточными медиаторами разнообразных функциональных возможностей мозга. Цепь обычно относится к набору взаимосвязанных компонентов, которые вместе выполняют определенную функцию. Нейронная цепь в мозге может представлять собой группу нейронов, получающих электрохимическую информацию, которую цепь модифицирует, и передает другим цепям для дальнейшей модификации. В качестве альтернативы нейронная цепь может состоять из сети взаимосвязанных областей мозга, которые вместе интегрируют огромные объемы информации и выполняют более сложные когнитивные и регуляторные функции. Ясно, что эти распределенные нейронные цепи не присутствуют ни при рождении, ни в течение жизни. На самом деле, развитие схемы мозга требует координации необычайно сложного набора событий, связанных с развитием нервной системы.
Структура и функции нервных цепей постоянно изменяются и развиваются с момента первого контакта между нервными клетками. Взаимодействие врожденных генетических программ с широким спектром воздействия окружающей среды и опыта определяет рождение, смерть и клеточные характеристики нейронов, а также формирование и преобразование их аксонов, дендритов и синапсов. Следовательно, нейронные цепи имеют разнообразные конфигурации и функциональные атрибуты в пределах конечного набора генетически и экологически ограниченного набора возможных конструкций и функций, в то же время в значительной степени разворачиваясь в соответствии с предсказуемой временной шкалой развития.
Гибкость и разнообразие результатов развития создают диалектику адаптации с уязвимостью при развитии от уровня отдельных контуров до взрослеющей личности. Возможности для адаптивных изменений и периоды критической уязвимости во время развития нейронных цепей сами по себе динамичны, специфичны во времени и чувствительны к опыту и воздействиям окружающей среды. По мере того как когнитивные функции, эмоции и поведение в норме и при болезни все чаще рассматриваются с точки зрения функционирования нейронных цепей, определение типичных траекторий созревания этих цепей создает важнейший фон, на котором генетические изменения и влияние неблагоприятного опыта на созревание мозга могут развиваться. сравнивать и противопоставлять, чтобы определить причины развития нервно-психических заболеваний.
Понимание процессов развития и организации функциональных свойств нейронных цепей требует доступа к определенным цепям с известными и измеримыми свойствами на разных этапах развития. Этические, методологические и практические соображения по понятным причинам ограничивают изучение зарождающейся нервной системы человека и содержащихся в ней нервных цепей (Huisman et al, 2002; Huttenlocher and Dabholkar, 1997; Levitt, 2003). Данные о развитии нервной системы человека и нечеловеческих приматов, состоящие в основном из патологоанатомических и нейровизуализационных исследований, ограничены и ограничены по своей природе (Fogliarini et al, 2005; Huttenlocher, 19).79, 1990; Хаттенлохер и Дабхолкар, 1997 г.; Левитт, 2003). Однако замечательная эволюционная консервативность событий развития нервной системы и их синхронизация у разных видов помогают подтвердить распространение знаний о пренатальном развитии мозга от животных к человеку (Bystron et al, 2008; Finlay and Darlington, 1995; Katz, 2007; Levitt, 2003; Лунд и Льюис, 1993; Марин-Падилья, 1988). В этом обзоре мы объединяем устоявшиеся и новые знания о развитии, чтобы описать нормальное созревание нейронных цепей. Мы также приводим конкретные примеры нервно-психических расстройств, которые обычно наблюдаются детскими психиатрами и детскими неврологами, чтобы проиллюстрировать, как знание нормального развития контуров может помочь в изучении и лечении психопатологий развития.
САМЫЕ РАННИЕ НЕРВНЫЕ ЦЕПИ
Ранние гестационные события создают основу для генеза нейронных цепей
Нейроонтогенный процесс у человека начинается в гестационном возрасте (ГВ) на 2-3 неделе со складыванием и слиянием эктодермы с образованием нервная трубка (Ladher and Schoenwolf, 2005) (рис. 1). На 4-й неделе беременности ростральная часть нервной трубки образует три пузырька, которым суждено дать начало переднему, среднему и заднему мозгам (Jessell and Sanes, 2000; Rash and Grove, 2006b; Rhinn et al, 2006). ; Стерн, 2001). Самый ростральный прозэнцефалический (передний мозг) пузырек затем образует два пузырька, которым суждено стать конечным мозгом (кора головного мозга) и промежуточным мозгом (таламус, гипоталамус и другие структуры). За этим следует сложная, динамичная, последовательная и тем не менее перекрывающаяся во времени серия клеточных событий, которые генетически детерминированы, эпигенетически направлены и находятся под влиянием окружающей среды. К 5-6 неделям ГС нейробласты или предшественники нейронов быстро пролиферируют в вентрикулярной зоне (зародышевый матрикс), которая выстилает желудочки головного мозга (Bystron et al, 2008; Ghashghaei et al, 2007; Hatten, 19).93; Джесселл и Санес, 2000 г.; Корнак и Ракич, 1995 г.; Левитт, 2003; Макманус и др., 2004 г.; Молливер и др., 1973; Монк и др., 2001; Пенсеа и др., 2001 г.; Ракич, 1978, 1982, 1988, 1995, 2003; Раш и Гроув, 2006а).
Хронология основных событий в развитии мозга. На этой диаграмме показано развитие мозга, начинающееся с нейрации и продолжающееся миграцией нейронов, синаптогенезом, обрезкой, миелинизацией и истончением коры. Воспроизведено с разрешения и изменено из Giedd (1999) (Авторское право 1999 г. ) Американская психиатрическая ассоциация.
Слайд PowerPoint
Изображение полного размера
Ламинарная структура коры головного мозга кодируется на ранних стадиях развития. К 8-й неделе гестации нейробласты начинают дифференцироваться либо в специфические типы нейрональных клеток, либо в макрогилу, в зависимости от их расположения в сложной топографической матрице молекулярных градиентов в слое желудочковой зоны (рис. 2). Постмитотические клетки мигрируют из этого слоя, образуя корковые пластинки, по принципу «изнутри наружу», при котором более глубокие слои коры формируются раньше, чем более поверхностные (Hatten, 19).93; Корнак и Ракич, 1995 г.; Ракич, 1978, 1988, 1995). Большинство постмитотических нейронов путешествуют по радиальным глиальным клеткам, которые служат проводниками на пути нейронов к их конечному пункту назначения (Rakic, 1972; Rakic et al, 1994b). Радиальная глия сама по себе может также давать начало нейронам в развивающейся коре (Liu and Rao, 2004; Malatesta et al, 2000; Miller, 2002; Miyata et al, 2001; Noctor et al, 2001). Миграция зависит от сложного набора молекулярных взаимодействий между нейронами и каркасной глией (Chao et al, 2009).; Грессенс, 2000; Хаттен, 1999). Другая меньшая группа нейронов берет начало из зачатков ядер базальных ганглиев (медиального и латерального ганглиозных возвышений) и мигрирует тангенциально (т. е. параллельно внешней поверхности коры) к местам назначения в развивающейся коре головного мозга и таламусе, давая начало всем нейронам. ГАМКергические нейроны в зрелом мозге (McManus et al, 2004; Monk et al, 2001; O’Rourke et al, 1992; Van Eden et al, 1989). Пик миграции нейронов приходится на 12-ю и 20-ю недели гестационного возраста и в основном завершается к 26-29 неделям гестационного возраста.(де Грааф-Питерс и Хаддерс-Альгра, 2006 г.; Гупта и др., 2005 г.) (рис. 2).
Эмбриональные слои развивающейся коры головного мозга человека. (а) Схематическая иллюстрация развития слоев неокортекса человека. ( б ) Гистохимический срез головного мозга плода человека в возрасте 16 недель, окрашенный крезиловым фиолетовым, чтобы показать расслоение коры. (a) Воспроизведено с разрешения Bystron et al (2008) (Copyright 2008) Nature Publishing Group; (b) воспроизведено с разрешения Kostović et al (2002) (Copyright 2002) Oxford University Press.
Слайд PowerPoint
Изображение в полный размер
Ошибки в миграции нейронов могут иметь серьезные последствия для развития нервной системы. Лиссэнцефалия, или «гладкий мозг», например, представляет собой нарушение миграции нейронов, которое нарушает нормальное формирование извилин и борозд. Его функциональные последствия варьируются от умственной отсталости до смерти в младенчестве (Olson and Walsh, 2002). Различные причины лиссэнцефалии включают мутации в генах, кодирующих белки цитоскелета, компоненты базальной мембраны, гликозилтрансферазы и компоненты рилин сигнальный путь. Нарушения миграции нейронов также могут вызывать очаги эктопической кортикальной ткани в белом веществе. Эти очаги серого вещества содержат как ГАМКергические, так и глутаматергические клетки, которые могут вызывать судорожные расстройства (Gomez et al, 1999; Uhlmann et al, 2002).
Ранние синапсы в развивающемся мозге
Когда нейроны завершают миграцию, их аксоны и дендриты направляются к соответствующим синаптическим партнерам. Каркасные клетки и молекулярные градиенты важны для сборки этих синаптических связей. Также присуще как построению, так и созреванию синапсов в нейронных цепях их постоянное уточнение и модификация. Синаптические связи между нейронами на ранних стадиях развития часто являются переходными ступеньками к более стабильным связям, которые характеризуют более зрелые цепи.
Самые ранние синаптические связи формируются примерно на 5-й неделе гестационного возраста нейронами, расположенными в первом распознаваемом слое коры, известном как препластинка (также называемая первичным плексиформным слоем) (Marin-Padilla, 1971; Raedler and Raedler, 1978; Rickmann et al. , 1977; Стюарт и Перлман, 1987; Супер и др., 1998; Вуд и др., 1992). На своем пути к предпластинке, а затем и к субпластинке, аксоны дорсальных таламических нейронов управляются молекулярными взаимодействиями с популяцией тангенциально мигрирующих нейронов, называемых «коридорными» клетками, которые, подобно клеткам радиальной глии, представляют собой класс каркасных клеток. клеток (Lopez-Bendito et al, 2006). Нейроны препластинки служат первоначальными синаптическими мишенями для нейронных проекций из развивающегося таламуса и ствола мозга. Клетки предпластинки удачно называют пионерными нейронами, потому что они образуют временные связи с пресинаптическими клетками, выступая в качестве мишеней-заполнителей до тех пор, пока соответствующие постсинаптические нейроны не будут готовы к формированию более зрелых связей (Chao et al, 2009).). Нейроны внутри препластины образуют примитивную, но функционально активную раннюю корковую цепь (Bayer and Altman, 1990; Kostovic and Molliver, 1974; Kostovic and Rakic, 1990; Marin-Padilla, 1971, 1978; Mrzljak et al, 1990).
Аксональный гликопротеин, молекула адгезии нервных клеток L1, опосредует взаимодействие между цитоскелетом и внеклеточным матриксом и, следовательно, играет важную роль в миграции и дифференцировке нейронов (Sonderegger and Rathjen, 1992; Thelen et al, 2002). Мутации в гене, кодирующем этот белок, вызывают Х-сцепленный синдром умственной отсталости CRASH (агенезия мозолистого тела, умственная отсталость, афазия, спастическая параплегия, гидроцефалия) (Jouet et al, 19). 94; Розенталь и др., 1992). Воздействие алкоголя нарушает функцию молекулы клеточной адгезии L1, что затем может способствовать проявлению алкогольного синдрома плода (Ramanathan et al, 1996; Tang et al, 2006).
Взлет и падение подпластины: от синапсов к ранним нейронным цепям
Подпластина представляет собой переходный, но важный эмбриональный кортикальный слой, который формируется внутри препластинки (Allendoerfer and Shatz, 1994; Bystron et al, 2008). Подпластинка толще всех других слоев коры между 18-й и 22-й неделями гестационного возраста, когда она в пять раз толще кортикальной пластинки. В этот период субпластинка богата синапсами и содержит признаки ламинарной организации. Дифференциальное время развития событий в разных областях мозга очевидно на субпластине, в которой соматосенсорные области развиваются раньше, чем зрительные области (Kostovic and Rakic, 19).90). Нейроны в этих частях субпластины получают предварительные афферентные импульсы от зрительного и соматосенсорного таламуса, холинергические афференты от базальных отделов переднего мозга и моноаминергические афференты от ствола мозга (Allendoerfer and Shatz, 1994; de Graaf-Peters and Hadders-Algra, 2006; Ghosh). et al, 1990; Ghosh and Shatz, 1992; Kostovic and Rakic, 1984, 1990; McConnell et al, 1989; Super et al, 1998).
Нейроны подпластины имеют молекулярные компоненты, необходимые для функциональной ГАМКергической передачи (Meinecke and Rakic, 1992), которая, в отличие от зрелой нервной системы, является возбуждающей на протяжении большей части раннего развития (Ben Ari et al, 1997). Электрофизиологические записи показали, что, хотя они электрически молчат в состоянии покоя, субпластинчатые нейроны способны распространять нервные сигналы через моноаминергические, холинергические и глутаматергические синапсы (Friauf et al, 1990; Isaac et al, 1997). Таким образом, архитектурная структура и функциональные возможности основных нейротрансмиттерных систем устанавливаются на ранних сроках беременности, даже несмотря на то, что их функция в раннем развитии коры у человека в настоящее время плохо изучена. Воздействие наркотиков или лекарств, которые изменяют передачу сигналов, метаболизм или другие элементы физиологии нейротрансмиттеров, может влиять на развитие нейронных цепей и систем нейромедиаторов глубоким или более тонким, но все же значительным образом.
Как и на препластинке, синаптические соединения в субпластине служат заполнителями перед более поздними, более устойчивыми соединениями для таламокортикальных нейронов. Таким образом, подкорковые афференты субпластины могут временно соединяться со своими будущими постсинаптическими мишенями в слое IV коры через промежуточные нейроны субпластины (Ghosh et al, 1990; Ghosh and Shatz, 1992), которые сами проецируются в слои будущей коры головного мозга. , корковая пластинка и маргинальная зона (Allendoerfer and Shatz, 1994; Канолд, 2004). После паузы в субпластине подкорковые афференты создают более постоянные связи внутри корковой пластинки посредством процесса синаптического уточнения, который медленно начинается примерно на 20-й неделе гестационного возраста, достигает своего пика между 24-й и 28-й неделями гестационного возраста и продолжается в перинатальный период. Это уточнение синаптических связей вызывает растворение субпластины, что можно наблюдать на МРТ плода (Huisman et al, 2002; Kostović et al, 2002). После 28 недель беременности нисходящая субпластинка в основном содержит нейроны, предназначенные для ассоциативных областей и комиссуральных путей, которые развиваются одними из последних корковых областей и путей (Kostovic and Rakic, 19).90).
РАЗВИТИЕ НЕРВНЫХ ЦЕПЕЙ В КОРТИКАЛЬНОЙ ПЛАСТИНКЕ В ПЕРИНАТАЛЬНОМ ПЕРИОДЕ
Обзор
По мере миграции постмитотических клеток на 7–8-й неделе ГА из желудочковой зоны в препластинку они формируют еще один зародышевый клеточный слой, известный как кортикальная пластинка (Bystron et al, 2008; Rickmann and Wolff, 1981; Super et al, 1998) (рис. 2). По мере того, как она формируется внутри препластинки, кортикальная пластинка разделяет препластинку на два слоя: субпластинку внизу и маргинальную зону вверху. Корковая пластинка будет формировать пластинки II–VI зрелой коры головного мозга, тогда как маргинальная зона в конечном итоге сформирует корковый слой I.
Растворение субпластины и одновременное созревание корковой пластинки сигнализирует о переходе в развитии корковых цепей. Временные и промежуточные связи теперь уступают место более устойчивым паттернам нейронных связей, которые обладают большим потенциалом для создания целенаправленной деятельности. По мере своего развития корковая пластинка все больше приобретает организационные черты зрелой коры. Ламинирование сначала присутствует в первичной сенсорной и моторной коре уже на 25-й неделе гестационного возраста. К 32-й неделе гестационного возраста развивающаяся кора имеет полный взрослый набор отчетливых вертикальных пластинок (Kostovic et al, 19).95), содержащие афференты всех основных нейротрансмиттерных систем (Levitt, 2003) и разнообразные типы дифференцированных клеток глии и нейронов (Lund and Lewis, 1993), включая возбуждающие глутаматергические шиповидные пирамидные нейроны и ГАМКергические непирамидные интернейроны (DeFelipe and Farinas, 1992; Ньювенхейс, 1994).
Ламинирование коры и локальные нейронные цепи
Подобно зрелой коре, развивающаяся корковая пластинка состоит из 3–6 клеточных слоев. В зависимости от конкретной области коры корковый слой I формируется между 24-й и 34-й неделями гестационного возраста, а слои III и IV появляются между 32-й и 34-й неделями гестационного возраста (Kostovic et al, 19).95). Каждый слой содержит определенный набор типов клеток, морфология и ламинарное расположение которых определяют паттерн локальных и отдаленных проекций, которые каждая клетка может отправлять или получать (рис. 3).
Разнообразие нейронов кортикальных пластинок. Реконструированный корональный вид различных типов клеток, представленных в зрелых слоях коры I–VI. Клетки были окрашены для удобства просмотра следующим образом: аксоны ярко-синего или ярко-желтого цвета; дендриты; темно-синий или темно-желтый. L, корковый слой; wm, белое вещество; б — корзинчатые клетки; db, ячейка с двойным букетом; р, пирамидальные клетки; ss — шиповатые звездчатые клетки. Масштабная линейка, 300 мкм. Воспроизведено с разрешения Binzegger et al (2004) (Copyright 2004) Society for Neuroscience.
Слайд PowerPoint
Изображение в натуральную величину
Клетки в соседней вертикальной пластинке организованы в функциональные радиальные онтогенетические столбцы, каждый из которых состоит из множества меньших миниколонок, самые длинные оси которых расположены перпендикулярно поверхности коры (Hubel, Wiesel, 1962; Mountcastle et al, 1957). ). Пирамидные нейроны клонально происходят из клеток радиальной глии и могут располагаться в нескольких слоях внутри одной и той же миниколонки (Yu et al, 2009).). По мере своего созревания, начиная примерно с конца третьего триместра, эти клонально полученные клетки предпочтительно формируют функциональные возбуждающие синаптические связи друг с другом, а не с соседними клетками, которые не произошли от одного и того же предшественника (Yu et al, 2009). Соединения ближнего действия связывают клетки близлежащих миниколонок по горизонтали и клетки разных пластинок внутри каждой колонки по вертикали (Mountcastle, 1997).
Различия между пластинками коры основаны на типах клеток и связях, которые преобладают в каждом слое. Канонический паттерн ламинарных связей, образующих корковые микросхемы, лучше всего выражен в зрительной коре (рис. 4). Слой VI содержит пирамидальные клетки с богатыми дендритными разветвлениями, которые проецируют возбуждающие аксоны к таламусу (Gilbert, 19).83; Марин-Падилья, 1970). Этот слой также посылает и получает возбуждающие и тормозные сигналы от более поверхностных слоев коры в каждой миниколонке. Он содержит локальные тормозные и возбуждающие интернейроны и получает входные данные от подкорковых и других слоев коры. Наличие и степень клеточной дифференцировки в слое IV отражают эволюционное происхождение и сложность кортикальной ткани на поверхности коры. Изокортикальные (также называемые гомотипическими или эуламиновыми) области содержат зернистый слой IV и обычно участвуют в процессинге более высокого порядка. Периаллокортальные (или переходные) области имеют менее развитый слой IV, тогда как аллокортикальные (или гетеротипические) области не имеют слоя IV и обычно представляют собой лимбическую кору. Из всех пластинок коры слой V содержит самые крупные пирамидные клетки, которые посылают возбуждающие отростки к базальным ганглиям, стволу мозга и спинному мозгу. Подобно слою VI и более поверхностным слоям, слой V также получает и посылает другие возбуждающие и тормозные проекции.
Схема связей между нейронами 3 и 6 слоев коры головного мозга у взрослых. Эта диаграмма представляет собой сводку данных парных внутриклеточных записей и заливки красителем связанных нейронов зрительной коры взрослых крыс и кошек. Треугольники, возбуждающие нейроны; круги, тормозные интернейроны. Ясно, что предстоит подробно описать еще много типов межпластинчатых соединений. Воспроизведено с разрешения (Thomson and Bannister, 2003) (Copyright 2003) Oxford University Press.
Слайд PowerPoint
Изображение полного размера
Возбудительные нейроны в слое IV, такие как пирамидные и шиповидные звездчатые клетки, являются первичными мишенями для таламических входов (Kostovic and Goldman-Rakic, 1983). Нейроны в слое III получают конвергентные входы от тормозных интернейронов, которые возникают в этом слое и в более глубоких слоях. Пирамидальные клетки III слоя проецируются на ипсилатеральные и контралатеральные области коры (Mrzljak et al, 19).88). Точно так же слой II получает разнообразные входные данные, но содержит меньшие пирамидальные нейроны, которые посылают проекции в менее отдаленные области коры. Наконец, связь и вычислительные функции самой поверхностной пластинки, слоя I, также наименее изучены. В слое I аксоны, исходящие из других областей коры, оканчиваются на апикальных дендритах пирамидных клеток, лежащих в более глубоких пластинках.
Хотя клеточный состав и паттерны связей этих корковых пластинок более гетерогенны, сложны и перекрываются, чем можно предположить из следующего схематического описания (Douglas and Martin, 2004; Gilbert, 1983; Thomson and Bannister, 2003), нейронные цепи, образованные связями с, от и внутри каждой пластинки зрительной коры, служат модельной областью для изучения развития микросхем в других областях коры (рис. 4). Например, в сильно упрощенной архетипической корковой микросхеме таламические афференты проецируются на слой IV, откуда возбуждающие нейроны проецируются на пирамидные нейроны слоев III и II. Возбуждающие пирамидные клетки в слое III проецируются на пирамидные клетки в слое V и в слое II. Пирамидные клетки в слое V проецируются на пирамидальные клетки в слоях III и IV. Наконец, пирамидные клетки слоя VI проецируются в слой IV.
Обычно ламинарные выходы также следуют характерной схеме. Слои II и III расширяют корковые афференты, при этом слой II выступает более локально, а слой III простирается на более отдаленные проекции. Слои V и VI проецируются на подкорковые области, слой VI проецируется в основном на таламус, а слой V проецируется на ствол мозга, средний мозг и базальные ганглии. Внутри каждой микросхемы тормозные интернейроны регулируют этот основной поток информации от одного возбуждающего нейрона к другому. Возбуждающие клетки в большинстве слоев проецируются на тормозные интернейроны в других слоях. В свою очередь тормозные интернейроны могут проецироваться на возбуждающие нейроны или на другие тормозные интернейроны. Таким образом, сложные связи развивающейся коры являются как вложенными, так и перекрывающимися, а их особенности сохранятся по мере их тонкой настройки на протяжении всей перинатальной и постнатальной жизни.
Прогрессивные и регрессивные силы формируют нейронные цепи
Динамика синаптогенеза в коре головного мозга развивающегося плода достаточно хорошо охарактеризована (Becker et al, 1984; Bourgeois, 1997; Bourgeois et al, 1994; Huttenlocher et al, 1982a; Michel and Garey, 1984; Mrzljak et al, 1988, 1992, 1990; Rakic et al, 1986). Плотность синапсов в корковой пластинке увеличивается со скоростью примерно 4% в неделю примерно до 26-28 недель ГС, что является временем пикового переноса афферентных синаптических связей с субпластины на корковую пластинку (Zecevic, 19).98). Разветвление дендритов и синаптогенез ускоряются в третьем триместре, вызывая утолщение развивающейся коры (Huttenlocher and Dabholkar, 1997), что совпадает с появлением корковых извилин и борозд на МРТ плода (Garel et al, 2001; Huisman et al, 2002). 34-я неделя GA знаменует начало пикового периода синаптогенеза, в течение которого каждую секунду образуется почти 40 000 новых синапсов, и этот процесс продолжается и в раннем постнатальном периоде жизни.
Время синаптогенеза различается в разных слоях коры и в развивающихся областях коры. Следуя секвенированию слоев коры изнутри наружу, синаптогенез начинается раньше в более глубоких слоях, чем в более поверхностных (Huttenlocher, 19).90). Хотя в нескольких исследованиях не сообщалось о различиях в скорости синаптогенеза в соматосенсорной, зрительной, префронтальной и лимбической коре (Bourgeois et al, 1994; Rakic et al, 1986; Zecevic, 1998; Zecevic et al, 1989), другие исследования показали, что синаптогенез начинается раньше в первичных двигательных областях и позже в передних областях, таких как префронтальная кора (ПФК) (Huttenlocher, 1984, 1990; Huttenlocher and Dabholkar, 1997).
Клеточные и молекулярные механизмы, лежащие в основе расширения развивающейся центральной нервной системы, уравновешиваются выбраковкой нейронов и их отростков (Cowan et al, 1984). Пик числа нейронов в мозге человека приходится на 28-ю неделю гестационного возраста. Однако до половины нейронов, образовавшихся в ходе нейрогенеза, погибают в процессе апоптоза или естественной гибели клеток к концу подросткового возраста (Lossi and Merighi, 2003). . Причины избыточной продукции большинства типов нейронов и глии до конца не изучены.
Многие из выбракованных нейронов элиминируются в течение двух дискретных эпох развития (Gohlke et al, 2006; Lossi and Merighi, 2003; Rakic and Zecevic, 2000). Первая волна апоптоза вовлекает в первую очередь пролиферирующие клетки-предшественники и молодые постмитотические нейробласты в желудочковой зоне (Blaschke et al, 19).96). Он начинается примерно на 7-й неделе гестационного возраста и продолжается в течение первого триместра. Эта волна управляется внутриклеточными процессами, связанными с регуляцией клеточного цикла (de la Rosa and de Pablo, 2000). Пик второй волны апоптоза приходится на 19–23 недели гестационного возраста и элиминирует постмитотические нейроны в кортикальной пластинке. В этой волне апоптоз регулируется синаптической активностью, различными формами межклеточных контактов и широким спектром диффундирующих и мембраносвязанных трофических факторов, которые продуцируются глией и др. нейронами (Lossi and Merighi, 2003). Клетки, которые посылают процессы в неправильные области, в конечном счете удаляются, потому что эти клетки с меньшей вероятностью активируются синхронно с другими входами в постсинаптическую клетку, которая затем не может высвобождать трофические факторы, необходимые для выживания пресинаптических нейронов (Eyre et al, 2001; Года). и Дэвис, 2003). Таким образом, когерентная активность нейронов также важна для выживания и поддержания нейронов, синапсов и нейронных цепей.
Глиальные клетки и миелинизация поддерживают развитие нейронных цепей
Микроглия, олигодендроциты и астроциты, глиальные компоненты белого вещества центральной нервной системы, участвуют во множестве важных вспомогательных функций нейронов, включая управление миграцией нейронов, регуляцию состава внеклеточной среды, модуляции синаптических связей и клиренса нейротрансмиттеров. Премиелинизирующие олигодендроциты заселяют развивающуюся кору во время беременности. Их последующая выработка миелина повышает скорость и точность передачи информации, закодированной в потенциалах действия, которые распространяются по нейронам. Между 20-й и 28-й неделями гестационного возраста зрелый миелин обнаруживается сначала в подкорковых областях, а затем в корковых областях. На 35-й неделе гестационного возраста он обнаруживается в прецентральной и постцентральной извилинах и зрительном луче, а на 40-й неделе гестационного возраста он присутствует в акустической радиации (Iai et al, 19).97). На МРТ в головном мозге недоношенных детей к 29-й неделе гестационного возраста можно различить некоторое количество миелинового белого вещества, но большая часть белого вещества выглядит немиелинизированной (Hüppi et al, 1998). Между 36-й и 40-й неделями гестационного возраста доля общего объема мозга, содержащего миелинизированное белое вещество, увеличивается с 1 до 5% (Hüppi et al, 1998).
Премиелинизирующие олигодендроциты могут быть особенно чувствительны к перинатальной гипоксии или ишемии, которые могут нарушать тракты белого вещества в лобных и височных долях, вызывая церебральный паралич или умственную отсталость (Back et al, 2001; Ness et al, 2001). Также считается, что воздействие на развивающийся мозг в это время токсинов окружающей среды, злоупотребления наркотиками, дефицита питательных веществ и последствий преждевременных родов нарушает миелинизацию и тем самым предрасполагает к плохим когнитивным, нервно-психическим и психическим последствиям.
Функциональные корреляты развития нейронных цепей в перинатальном периоде
Возрастающая организационная сложность и зависящее от активности ремоделирование нейронных связей создают структурную основу, поддерживающую ранние функциональные свойства корковых цепей. Нейроны с множественными ветвями аксонов могут иннервировать и влиять на многочисленные клетки-мишени, тогда как сложные дендритные деревья позволяют синтезировать входы от других клеток, которые многочисленны и разнообразны (Jessell and Sanes, 2000). Оттачивание связности между этими морфологически и нейрохимически разнообразными популяциями нейронов порождает соответственно различные вычислительные функции, варьирующиеся от ограниченного ввода и отклика до сложной интеграции множества сигналов, которые вместе усваивают информацию из разных областей мозга (де Грааф-Питерс и Хаддерс). -Algra, 2006; Джесселл и Санес, 2000; Левитт, 2003). Таким образом, повышение структурной организации предвещает повышение функциональной организации корковых цепей.
Исследование функциональной активности головного мозга плода in vivo является сложной задачей. Тем не менее, электрофизиологические измерения у недоношенных детей показали, что модели электрической активности меняются параллельно с анатомическим развитием коры между 20 и 40 неделями гестационного возраста (Graziani et al, 1968; Kostovic et al, 1995; Kurtzberg et al, 1984; Novak). и др., 1989 г.; Воан и Курцберг, 1989 г.). Эти связанные с развитием изменения в сигнале ЭЭГ, наиболее заметные в сенсомоторной коре, могут отражать повышенную активность нейронов поверхностного коркового слоя III по сравнению с более глубоким корковым слоем IV (Kostovic et al, 19). 95). Поскольку слой IV получает таламический вход, тогда как слой III проецируется на другие слои коры и, следовательно, поддерживает обработку более высокого порядка, данные ЭЭГ о повышенной активности в слое III могут отражать функциональное созревание сенсомоторной коры. Эти ранние события созревания предвещают последующее более широкое функциональное развитие как локальных, так и распределенных корковых цепей.
РАЗВИТИЕ НЕРВНЫХ ЦЕПЕЙ В МЛАДЕНЧЕСТВЕ
Обзор
Ранний постнатальный период представляет собой время кардинальных изменений в структуре и функциях мозга. Мозг вырастает примерно до 70 % своего взрослого размера к 1 году и примерно до 80 % взрослого размера к 2 годам (Декабан, 19 лет).78; Никмейер и др., 2008). Это увеличение объема мозга в течение первого года жизни является наибольшим в мозжечке, за которым следуют подкорковые области, а затем кора головного мозга (Knickmeyer et al, 2008), которая увеличивается в объеме на впечатляющие 88% в первый год жизни, 15% на второй год, а затем более скромно, но стабильно (Knickmeyer et al, 2008). Развитие серого и белого вещества, миелинизация, синаптогенез, обрезка и синаптическая модификация устанавливают фундаментальную анатомическую организацию для начального функционирования нейронных цепей внутриутробно . Эти прогрессивные и регрессивные процессы развития продолжаются в ранней постнатальной жизни. Впоследствии локальные связи внутри корковых цепей точно настраиваются, а сложные связи дальнего действия между цепями формируют все более унифицированную и функционально организованную нейронную сеть.
В раннем постнатальном периоде новорожденный не просто пассивно испытывает новые стимулы; младенец также вырабатывает модели поведения, которые сильно влияют на его окружение (Kandel et al, 2000). Вместе с врожденными генетическими факторами новые эмпирические входные данные и поведенческие реакции воздействуют на еще незрелый мозговой субстрат, стимулируя дальнейшее развитие нейронных цепей. Со временем опыт играет все более заметную роль в формировании нейронных цепей.
Раннее постнатальное развитие нейронных цепей
Мозг новорожденного в возрасте 2–4 недель примерно на 36 % превышает размер мозга взрослого человека (Knickmeyer et al, 2008). Гистологическое исследование головного мозга новорожденных показало, что зернистый слой IV уже присутствует во всех областях неокортекса (Kostovic, Rakic, 1980). Первичные области коры, включая моторную, соматосенсорную, зрительную и слуховую кору, могут быть идентифицированы цитоархитектонически. Ассоциативная кора, однако, в этом возрасте менее четко очерчена.
Хотя макроскопическая визуализация структуры головного мозга с помощью МРТ у недоношенных и доношенных детей не может дать нам информации о клеточных процессах нормального развития мозга, нейровизуализация дает редкое представление об анатомических особенностях развития мозга на поздних сроках беременности и в раннем постнатальном периоде. Анатомическая МРТ недоношенных детей показывает, что общий объем головного мозга увеличивается почти в три раза между 29-й и 41-й неделями гестации (Hüppi et al, 1998). В этот период серое вещество разрастается от 35 до 50% всего объема мозга. МРТ доношенных детей в возрасте 39 лет-48 недель показывает сильное увеличение общего объема серого вещества в этом окне развития по сравнению с другими отделами мозга (Gilmore et al, 2007b). У доношенных детей серого вещества больше в сенсомоторной и зрительной коре, а также в теменно-затылочной области по сравнению с недоношенными детьми (Peterson et al, 2003). Рост серого вещества выражен в затылочной и теменной коре в первые недели жизни.
В раннем постнатальном периоде кортико-кортикальные волокна в остатках субпластины соединяются со своими корковыми постсинаптическими мишенями (Kostovic and Rakic, 19).90; Шварц и др., 1991). Клетки, экспрессирующие маркеры, характерные для нейробластов и ранних нейронов, идентифицированы в субвентрикулярной зоне новорожденных (Weickert et al, 2000), а мигрирующие нейроны все еще можно обнаружить в областях белого вещества (Kostovic and Rakic, 1980). Хотя корковый нейрогенез и миграция нейронов продолжаются в постнатальный период (Bhardwaj et al, 2006; Shankle et al, 1999), более вероятно, что экспансия глии и миелинизация в этот период вносят более значительный вклад в резкий рост мозга, наблюдаемый у новорожденных и малышей (Декабан, 1978). Таким образом, перинатальное и раннее постнатальное развитие мозга характеризуется началом миелинизации и поразительным развитием связей серого вещества, особенно в сенсомоторной и зрительной коре.
Аномальный размер мозга в раннем постнатальном периоде часто предвещает более позднее начало нарушений развития нервной системы. Недоедание, например, уменьшает размер мозга в младенчестве и оказывает широкий спектр побочных эффектов на развитие нервной системы (Holden, 2007). И наоборот, ускоренный рост мозга в младенчестве, по-видимому, является одним из ранних проявлений широкого спектра задержек развития двигательных, языковых и когнитивных функций, включая аутизм (Courchesne et al, 2001; Hazlett et al, 2005). Результаты МРТ у аутичных людей указывают на расширение компартментов как серого, так и белого вещества (Hardan et al, 2006; Piven et al, 19).95). Эти находки могут указывать на персистенцию избыточных синапсов и миелина или на сниженный апоптоз, а также на аномальные паттерны связей мозга (Belmonte et al, 2004). Однако точные клеточные и молекулярные детерминанты увеличения размера мозга у детей с аутизмом еще предстоит определить.
Синаптогенез и сокращение синапсов и аксонов в младенчестве
Корковое серое вещество продолжает расти в течение первых нескольких лет жизни, хотя его скорость наиболее высока в течение первого постнатального года. Траектория расширения серого вещества была показана с помощью МРТ (Knickmeyer et al, 2008) и согласуется с посмертными исследованиями, показывающими, что развитие дендритов, шипов и синапсов продолжает ускоряться с почти логарифмической скоростью в течение первых 350–400 лет. послеродовых дней (Андерсен, 2003; Буржуа, 19 лет).97; Лунд и Льюис, 1993 г.; Супер и др., 1998). В первые 2 года жизни наблюдается разветвление как пирамидных клеток, так и ГАМКергических тормозных интернейронов (Mrzljak et al, 1990), а также относительное расширение слоев коры II и III по сравнению с другими слоями (Huttenlocher and Dabholkar, 1997; Landing и др., 2002; Зечевич и др. , 1989).
Временной ход синаптогенеза различается в разных областях коры. В первичной зрительной коре, например, после всплеска образования синапсов в возрасте от 3 до 4 месяцев плотность синапсов достигает своего пика на уровне 140–150% взрослого уровня в возрасте от 4 до 12 месяцев, после чего среднее число количество синапсов на один нейрон снижается (Буржуа, 1997; Буржуа и Ракич, 1993; Хаттенлохер, 1990; Huttenlocher и др., 1982а, 1982b; Мишель и Гэри, 1984). Синаптогенез в префронтальной коре начинается примерно в то же время, что и в зрительной коре, но не достигает своего пика до возраста 8 месяцев, продолжаясь после этого в течение второго года жизни (Huttenlocher and Dabholkar, 1997; Kostovic et al, 1995).
Уничтожение аксонов, дендритов и синапсов, а также гибель нейронов в результате апоптоза являются важными параллельными процессами образования дополнительных аксонов, дендритов и синапсов (Cowan et al, 19).84). Процессы обрезки начинаются на поздних сроках беременности и становятся все более активными после рождения. Как и в случае с синаптогенезом, временной ход обрезки различается в разных областях мозга: сенсорная и моторная кора претерпевает драматическую тонкую настройку после рождения, за ней следуют ассоциативная кора и мозолистое тело, а затем области, которые обеспечивают высшие когнитивные функции (Levitt, 2003). .
Пластичность синапсов и развитие нейронных цепей в постнатальной жизни
Подобно сокращению, ремоделирование существующих синапсов играет ключевую роль в реорганизации и тонкой настройке нейронных цепей. Синаптическая пластичность относится к набору механизмов, которые опосредуют зависящее от активности усиление или ослабление нейронных связей на уровне синапса (Citri and Malenka, 2008). Сила и характер активности в данном синапсе вызывают временную или длительную депрессию или усиление связи между нейронами (Martin et al, 2000; Morris, 2006; Squire et al, 2004). Таким образом, длительная потенциация и длительная депрессия являются активными процессами, зависящими от электрической активности. При синдроме ломкой X наиболее распространенной генетической причиной умственной отсталости является замалчивание 9Ген 0083 Fmr1 (Devys et al, 1992), по-видимому, нарушает баланс между долговременной потенциацией и депрессией (Bagni and Greenough, 2005; Huber et al, 2002). Широкие эффекты нарушений в зависящей от активности модификации синапсов у Fragile X подчеркивают критическую важность молекулярных и клеточных механизмов, лежащих в основе синаптической пластичности, для нормального постнатального развития мозга и здоровых когнитивных результатов.
Реорганизация таламокортикальных проекций путем обрезки и ремоделирования синапсов также имеет решающее значение для функции ряда сенсорных систем. Например, при развитии бинокулярного зрения правильная дифференциация столбцов в зрительной коре зависит от долговременной потенциации, долговременной депрессии и сокращения, которые регулируются паттернами нейронной активности, возникающими в обеих сетчатках (Берарди). и др., 2000; Эрлих и Малинов, 2004; Хаттенлохер и др. , 1982б; Такахаши и др., 2003). В соответствии с моделью обучения, основанной на Хеббе, конкуренция между скоординированными действиями, возникающими в сетчатке, приводит к поэтапному выбору входных данных от одного глаза для каждого нейрона. Итеративная скоординированная активность стабилизирует синаптические связи от одного глаза, тогда как синапсы с контралатеральными афферентами, которые передают более слабые и менее организованные паттерны активности, становятся менее активными и ликвидируются (Hooks and Chen, 2006).
Поскольку нейронная активность смягчает важные влияния опыта и окружающей среды, которые формируют развитие нейронных цепей, изменения в зрительных сенсорных входных данных и, таким образом, неупорядоченный паттерн нейронной активности в этот период развития вызывают относительно непоправимые нарушения зрения (Huberman, 2007). ). Например, неспособность исправить врожденную катаракту у младенцев к 6-месячному возрасту или косоглазие к 7-летнему возрасту приводит к необратимым нарушениям в зрительной системе (Levi and Li, 2009). ; Маурер и др., 1999). Таким образом, определенные эпохи в созревании нейронных цепей для зрения, по-видимому, представляют собой критические периоды уязвимости развития, периоды, когда эмпирический ввод необходим для нормального развития конкретных нейронных цепей и их функциональных возможностей и без которого потенциал для развития этих функциональных возможности теряются навсегда (Morishita and Hensch, 2008).
Миелинизация и развитие нервной системы
Вскоре после рождения глиальные клетки резко увеличиваются в размере и количестве, а скорость миелинизации продолжает пренатальную траекторию. Миелинизация протекает быстро в течение первого года жизни, а затем продолжается более медленными, но устойчивыми темпами (Barnea-Goraly et al, 2005; Gao et al, 2009).). Подобно синаптогенезу и сокращению, миелинизация имеет специфический временной ход, который различается в разных областях коры и с разной скоростью в определенных функциональных цепях. Посмертные и анатомические МРТ-исследования показывают, что миелинизация прогрессирует в направлении назад-вперед, следуя общей схеме созревания нервных цепей (Brody et al, 1987; Girard et al, 1991; Kinney et al, 1988; Paus et al, 2001). Первыми миелинизируются сенсорные пути, затем моторные пути, а затем ассоциативные области. Миелинизация достигает самых дальних участков лобных долей в возрасте от 7 до 11 месяцев (Баркович и др., 19).88). Внутри данной функциональной цепи подкорковые структуры миелинизируются раньше корковых областей.
Миелинизация, наблюдаемая с помощью анатомической МРТ, может отставать от ультраструктурных признаков миелинизации, особенно в возрасте до 3 лет (Matsuzawa et al, 2001; Paus et al, 2001). Диффузионно-тензорная визуализация (DTI) измеряет ограничения на диффузию воды вдоль пучков нервных волокон, в которой преобладает увеличение концентрации миелина во время развития белого вещества, и может обнаруживать более ранние признаки миелинизации (Bansal et al, 2007). Результаты DTI показали временной ход созревания белого вещества мозга, который в целом согласуется с результатами других экспериментальных модальностей (Berman et al, 2005; Gao et al, 2009).; Гилмор и др., 2007а; Hermoye и др. , 2006 г.; Макгроу и др., 2002 г.; Мукерджи и др., 2001 г.; Партридж и др., 2005 г.; Zhai et al, 2003) (рис. 5). Подобно DTI, результаты протонной магнитно-резонансной спектроскопии (MRS) развития подтверждают наблюдения, сделанные с использованием других экспериментальных подходов, которые показывают первоначальную быструю разработку нейронных процессов и синаптогенеза в первые 2 года жизни, за которой следует последующее плато (Horska et al, 2002). ; Kreis et al, 1993; Toft et al, 1994), которая различается по времени в разных областях коры (Cady et al, 19).96; Ханаока и др., 1998 г.; ван дер Кнаап и др., 1990).
Миелинизация мозга в процессе развития. Верхние панели: Т1-взвешенные аксиальные МРТ-изображения, полученные в продольном направлении от одного ребенка, демонстрирующие возрастное увеличение размера мозга и интенсивности белого вещества. Нижние панели: DTI-изображения трактографии белого вещества в поперечном сравнении, показывающие возрастное увеличение организации белого вещества мозолистого тела. Каждая панель представляет один предмет. Цветовая шкала представляет фракционную анизотропию; более высокие значения коррелируют с большей организацией дорожек волокон. Панели помечены возрастом участников на момент сканирования. А, передний; Л ушел. Воспроизведено с разрешения Gilmore et al (2006) (Copyright 2006) Американской психиатрической ассоциации.
Слайд PowerPoint
Изображение в натуральную величину
Функциональные визуализирующие исследования головного мозга младенцев: метаболическая активность в младенчестве, измеренная с помощью ПЭТ
Поскольку они включают воздействие радионуклеотидов и являются относительно инвазивными методами, позитронно-эмиссионная томография (ПЭТ) визуализирующих исследований здоровых детей является необычной. ПЭТ-визуализация небольшой клинической популяции с почти нормальным неврологическим обследованием (аномалии включали дистресс плода, судороги, врожденный порок сердца и синдром Стерджа-Веббера) показала, что подкорковые области и сенсомоторная кора являются наиболее метаболически активными областями мозга у новорожденных младшего возраста. старше 5 недель (Чугани, 19 лет)94; Чугани и др., 1987). К 3-месячному возрасту метаболическая активность увеличивается в теменной, височной и дорсолатеральной затылочной коре. К 6–8-месячному возрасту метаболическая активность лобных областей начинает увеличиваться (Chugani, 1994; Chugani et al, 1987). Подобно другим методам нейровизуализации, ПЭТ показывает, что модель изменения метаболической активности согласуется с вышеупомянутой пространственной изменчивостью во времени миелинизации и синаптогенеза.
Развитие функциональной связности в младенчестве
Развитие функциональной визуализации включает использование измерений мозговой активности, полученных в состоянии покоя, для выявления функционально значимых паттернов активности, которые представляют внутреннюю спонтанную мозговую активность (Binder et al, 1999; Biswal et al, 1995; Hampson et al, 2002; Мазоер и др., 2001). Считается, что наборы областей, которые имеют коррелированную во времени активность в состоянии покоя, составляют функциональные сети (Beckmann et al, 2005; Damoiseaux et al, 2006; Deluca et al, 2006; Fox et al, 2005, 2006; Fox and Raichle, 2007; Greicius). и др., 2003; Келли и др., 2008; Райхл и др., 2001). Эти методы показали ряд различных функциональных сетей у новорожденных в возрасте 39 лет.–44 недели, включая зрительные, сенсомоторные и слуховые сети обработки, а также префронтальную сеть (Fransson et al, 2007). Кроме того, функционально коррелированные медиальные и латеральные теменные области могут представлять собой рудиментарную сеть пассивного режима, систему, которая в зрелом возрасте состоит из предклинья, срединной префронтальной и латеральной теменной областей и которая связана с абстрактной и автобиографической самофокусированной обработкой информации (Gusnard). и др., 2001; Уддин и др., 2007). Это предполагаемое присутствие сети в режиме по умолчанию согласуется с результатами ERP, предполагая, что межполушарные связи возникают раньше, чем переднезадние связи дальнего действия (Bell and Fox, 19).92). Методы функциональной связи также предполагают, что сенсорные сети могут быстро развиваться в первый год жизни, опережая аналогичные изменения в зрительной сети, которые более заметны на втором году жизни (Lin et al, 2008).
Поведенческие корреляты развития нейронных цепей в младенчестве
Хотя развитие широкого спектра различных функциональных способностей, таких как двигательные, сенсорные и когнитивные функции, широко описано, методологические ограничения в изучении функции мозга в младенчестве ограничивают наши исследования. понимание развития конкретных нейронных цепей, которые обслуживают эти способности. Новорожденные демонстрируют сложные, самопроизвольные, но дезорганизованные движения, которые не кажутся целенаправленными (de Vries et al, 19).85; Прехтл и Хопкинс, 1986). Саккады глаз в этом возрасте вызываются рефлекторно и направляются стимулами окружающей среды, а не эндогенным контролем (Atkinson, 1984), что соответствует созреванию соматосенсорных и моторных областей перед зрительной и ассоциативной корой. Поведенческие изменения, которые отмечаются на третьем месяце жизни, в том числе тормозной контроль над рефлекторным поведением и саккадами, а также целенаправленное поведение, такое как целенаправленная координация головы и глаза и стремление схватить, могут отражать ремоделирование коры и миелинизацию ассоциативных областей. происходящие в то время (Гудкин, 1980). Появление передачи из рук в руки примерно в 6-месячном возрасте также соответствует по времени усилению анатомической и функциональной координации между сенсорными, моторными и ассоциативными цепями (Rothbart et al, 1990).
Уязвимость развивающейся нервной системы в младенчестве
Мозг чувствителен к вредным воздействиям, таким как токсины, наркотики, дефицит питательных веществ, инфекции, медицинские заболевания и стрессовые факторы окружающей среды на протяжении всей жизни. Однако, поскольку определенные процессы развития нервной системы происходят в относительно узких временных окнах, созревающий мозг во время внутриутробной жизни и в младенчестве может быть особенно восприимчив к неблагоприятным воздействиям. Например, ранняя разлука с опекунами, жестокое обращение, пренебрежение или социальная депривация в младенчестве или раннем детстве могут привести к стойкому поведенческому и нейрокогнитивному дефициту (Carpenter and Stacks, 2009). ; Креппнер и др., 2007). И наоборот, ранние вмешательства, направленные на устранение или ослабление последствий этих воздействий в конкретный период развития, могут предотвратить негативные последствия (Tarabulsy et al, 2008; Welsh et al, 2007). Исследования на людях, основанные на животных моделях, показали, что взаимодействие раннего жизненного опыта (например, стресса, социализации и материнской заботы) с генетическими вариациями в промоторе переносчика серотонина влияет на фенотипическое выражение поведенческих черт и психопатологий (Caspi et al., 2003; Угер и Макгаффин, 2008). Точно так же исследования на животных показали, что материнская забота влияет на поведение и будущее воспитание потомства посредством эпигенетической модификации хроматина в определенных генетических локусах (Francis et al, 19).99). Тем не менее, критические или чувствительные периоды развития уязвимости к развитию поведенческих черт и психопатологии не были определены так четко, как они были определены для развития сенсорных и моторных систем.
РАЗВИТИЕ НЕРВНОЙ ЦЕПИ В ДЕТСТВЕ И ПОДРОСТКОВОСТИ
Обзор
Хотя размер мозга увеличивается только с 80 до 90% от его взрослого размера в возрасте от 2 до 5 лет (Dekaban, 1978), миелинизация и ремоделирование синапсов особенно активны во время это так называемая фаза «плато» в развитии. Наше понимание клеточных изменений на этом этапе развития отстает от знаний о вычислительных возможностях мозга, полученных в результате поведенческих исследований у детей (Даймонд, 19 лет).95). Точно так же растущий объем продольных данных нейровизуализации описывает возрастные изменения морфологических особенностей и функциональной организации мозга в детстве и подростковом возрасте, хотя точные корреляты этих наблюдений на клеточном и молекулярном уровне в значительной степени спекулятивны.
Развитие нервной цепи в раннем и среднем детстве
Гистологические исследования показывают, что синаптогенез продолжается после младенчества, но более медленными темпами, поскольку серое вещество коры продолжает увеличиваться в объеме в возрасте 4–5 лет (Levitt, 2003). Как и в другие эпохи развития, динамика ремоделирования синапсов различается в разных областях коры. Синаптическая плотность достигает пика сначала в первичных сенсорных областях, затем в ассоциативных областях, а затем в префронтальной коре, которая отвечает за высшие когнитивные функции. Точно так же локальные корковые связи появляются раньше более отдаленных. Утолщению коркового слоя II, который создает короткие корковые связи с другими областями коры, предшествует утолщение слоя III, который создает более длинные связи с другими областями коры, в возрасте от 15 до 72 месяцев (Landing et al, 2002). В префронтальной коре абсолютное число дендритных шипиков на пирамидальных нейронах слоя IIIc, являющихся основным источником ассоциативных и комиссуральных проекций, достигает максимума при 29в месяц и снижается после 6 лет (Kostovic et al, 1995).
Чистое устойчивое состояние количества и плотности кортикальных синапсов, которое характеризует фазу плато развития, как полагают, отражает уравновешивающие эффекты увеличения скорости сокращения синапсов и замедления образования синапсов (Levitt, 2003). Поперечные исследования ПЭТ показывают, что к концу первого года жизни метаболизм глюкозы в головном мозге становится качественно сходным с таковым у взрослых (Chugani, 19).94; Чугани и Фелпс, 1986 г.; Чугани и др., 1987). Однако к 4–5 годам общий метаболизм мозга повышается в два раза по сравнению со взрослыми и остается высоким до 9–10 лет. Повышенная метаболическая активность мозга, вероятно, отражает бурный рост нейронов и глии в позднем младенчестве и раннем детстве. Устойчиво высокие уровни метаболической активности в течение периода плато могут отражать энергетические потребности миелинизации и ремоделирования синапсов, а также сокращения.
Миелинизация нейронных цепей в детском и подростковом возрасте
Исследования нейровизуализации внесли значительный вклад в наше понимание изменений серого и белого вещества в процессе созревания в ходе нормального развития мозга. Объемы белого вещества продолжают увеличиваться в детстве, подростковом и взрослом возрасте (Bartzokis et al, 2003; Benes et al, 1994; Courchesne et al, 2000; Giedd et al, 1999; Matsuzawa et al, 2001; Paus et al, 2001; Соуэлл и др. , 2003а). Скорость увеличения объемов белого вещества наибольшая в младенчестве. Рост в этом отделе продолжается более медленными темпами в детстве, подростковом и взрослом возрасте и достигает пика в середине пятого десятилетия жизни (Matsuzawa et al, 2001; Paus et al, 2001). Исследования DTI показывают, что темпы увеличения показателей организации белого вещества максимальны в возрасте до 10 лет (Ashtari et al, 2007; Barnea-Goraly et al, 2005; Ben Bashat et al, 2005; Giorgio et al, 2008; Huang et al. , 2006; Клингберг и др., 1999; Шмитхорст и др., 2002, 2005; Снук и др., 2005). Организация белого вещества происходит с разной скоростью между областями белого вещества и отдельными трактами белого вещества и продолжается и во взрослом возрасте.
Траектории развития коры головного мозга в позднем детстве и подростковом возрасте: данные анатомической визуализации детском или подростковом возрасте (рис. 6) (Giedd et al., 19 лет).99, 1996а, 1996б; Гогтай и др., 2004 г.; Джерниган и др.
, 1991; Ленрут и др., 2007 г.; О’Доннелл и др., 2005 г.; Пфеффербаум и др., 1994; Шоу и др., 2008 г.; Соуэлл и др., 2003a, 2004, 2007; Томпсон и др., 2005). Истончение коры происходит в направлении «сзади вперед» и происходит сначала в сенсомоторных областях, затем в ассоциативных областях и, наконец, в областях коры более высокого порядка, таких как верхняя префронтальная кора и задняя теменная кора (см. loni.ucla.edu/~thompson/DEVEL/dynamic.html для покадровых изображений истончения коры). Рисунок 6 Созревание серого вещества в процессе развития. Виды мозга справа и сверху, показывающие динамическую последовательность временных изменений объема серого вещества на поверхности коры. Изображения представляют смоделированные данные 52 анатомических МРТ-сканирований 13 человек в возрасте от 4 до 21 года, каждое из которых сканировалось четыре раза с интервалом примерно в 2 года. Цветовая шкала представляет единицы объема серого вещества. Воспроизведено с разрешения Gogtay et al (2004) (Copyright 2004) Национальной академии наук США.
Слайд PowerPoint
Полноразмерное изображение
Сообщалось также о локальном возрастном увеличении толщины коры головного мозга. Толщина перисильвиевой коры в нижней теменной и задней височной областях и особенно в левом полушарии, по-видимому, увеличивается между детством и взрослым (Sowell et al, 1999, 2001, 2003a). Это региональное утолщение сменяется на пятом десятилетии жизни истончением, совпадающим по времени с периодом замедленного истончения более дорсальных областей коры. Параллельно с изменениями толщины коры региональный рост, который, как считается, отражает увеличение объемов белого вещества, наблюдается в большей части префронтальной коры, ростральной передней части поясной извилины и височных полюсах (Sowell et al, 2003a). Сообщалось также о возрастных изменениях объемов подкоркового серого вещества с наблюдаемым уменьшением серого вещества базальных ганглиев (особенно хвостатого ядра) и увеличением структур медиальной височной доли, таких как миндалевидное тело и гиппокамп.
Эти связанные с развитием изменения толщины коры и объема мозга согласуются с нашим пониманием сроков созревания когнитивных функций, таких как внимание, рабочая память, когнитивный контроль и подавление реакции. Дальнейший анализ продольных анатомических данных МРТ выявил ряд функциональных коррелятов созревания коры. В одном отчете предполагается, что скорость изменения толщины коры в областях, отвечающих за когнитивные функции более высокого порядка (таких как вентролатеральная префронтальная, верхняя префронтальная и задняя теменная кора), коррелирует с показателями интеллекта (Shaw et al, 2006). В других сообщениях предполагается, что истончение коры в моторных областях коррелирует с мелкой моторикой, а утолщение коры в речевых областях левого полушария может коррелировать с навыками фонологической обработки (Lu et al, 2007; Sowell et al, 2004). Хотя истончение коры рассматривается как надежный маркер созревания, конкретные когнитивные корреляты этих анатомических изменений остаются неясными.
При шизофрении, начинающейся в детстве, корковое созревание происходит в целом по нормальному типу истончения, но оно происходит более быстрыми темпами и занимает более обширное пространство, чем у типично развивающейся молодежи (Thompson et al, 2001). Чрезмерное истончение сначала обнаруживается в теменной коре, которая служит зрительно-пространственной и ассоциативной обработке, а затем распространяется на височную и префронтальную кору. Степень потери серого вещества, по-видимому, коррелирует с тяжестью психотических симптомов. Считается, что это уменьшение ткани коры связано с уменьшением разветвления дендритов и уменьшением количества клеток, включая глию и сосудисто-нервную систему (Selemon and Goldman-Rakic, 19).99), но чаще всего они рассматриваются как следствие чрезмерного сокращения синапсов или аномальной синаптической пластичности (Feinberg, 1982; Liang et al, 2006; Micheloyannis et al, 2006; Rapoport and Gogtay, 2008). Наоборот, данные продольной анатомической МРТ при синдроме дефицита внимания с гиперактивностью (СДВГ) выявляют задержки во времени созревания коры, при этом пиковая толщина коры достигается позже, чем у типично развивающихся детей, особенно в передней височной и префронтальной коре (Shaw et al, 2007). ).
Предполагаемые механизмы изменения толщины серого вещества
Клеточная основа изменений толщины коры еще не установлена. Однако многочисленные доказательства привели к консенсусной гипотезе о том, что истончение коры может представлять собой два параллельных процесса: обрезку и миелинизацию. Считается, что вместе они улучшают возможности подключения в локальных и распределенных сетях, а также повышают эффективность и точность передачи сигналов. Следовательно, истончение ассоциативной коры все чаще рассматривается как маркер созревания коры у здоровых детей.
В соответствии с этим мнением исследования на животных и на людях показали возрастное снижение плотности синапсов в возрасте от 2 до 16 лет (Bourgeois and Rakic, 1993; Huttenlocher, 1979; Huttenlocher and Dabholkar, 1997; Rakic et al, 1986). . У мышей нейропиль, или аксональные и дендритные отростки, составляет примерно 60% объема коры (Bourgeois and Rakic, 1993), и обрезка этих отростков может представлять собой источник истончения коры, обнаруженный с помощью МРТ (Paus et al, 2008). Гипотеза о том, что истончение коры представляет собой обрезку синапсов, аксонов и дендритов, также подтверждается данными о том, что общий метаболизм мозга, который в значительной степени зависит от потенциалов мембран нейронов в состоянии покоя (Mata et al, 1980; Nudo and Masterton, 1986), начинает снижаться в возрасте 9–10 лет и достигает взрослого уровня к 16–18 годам (Chugani, 1994; Chugani and Phelps, 1986; Chugani et al, 1987). Следовательно, сокращение синаптических процессов может связывать истончение коры с возрастным снижением метаболизма мозга (Mata et al, 1980; Nudo and Masterton, 1986). Миелинизация интракортикальных аксонов и увеличение объема белого вещества по отношению к объему серого вещества также могут влиять на количественную оценку серого вещества по данным МРТ (Paus, 2005; Paus et al, 2008; Sowell et al, 2003a; Yakovlev and Lecours). , 1967). Эта возможность подтверждается данными о том, что региональный рост размера мозга, который, как считается, отражает увеличение объема белого вещества, происходит в сочетании с истончением коры (Sowell et al, 2003a).
Хотя генетические факторы в значительной степени способствуют развитию и созреванию нейронных цепей, большое количество данных показывает, что тонкая настройка нейронных цепей также находится под влиянием окружающей среды. Исследования на людях показывают, что зависимая от активности элиминация возбуждающих связей в основном ответственна за 40-процентное сокращение синапсов в подростковом возрасте (Huttenlocher, 19).84; Ракич и др., 1994а). Напротив, уровни местных ГАМКергических тормозных интернейронов остаются довольно стабильными с детства до зрелого возраста (Changeux and Danchin, 1976; Innocenti, 1981). Взаимосвязь между истончением коры, миелинизацией и зависимым от опыта ремоделированием нейронных цепей становится все более ясной, но многое еще предстоит сделать для картирования макро- и микрохарактеристик этих процессов, а также для понимания их функциональных коррелятов.
Изменения в организации нейронных цепей в ходе развития: данные визуализирующих измерений функциональной связи
Корреляционный анализ данных фМРТ, полученных в состоянии покоя, недавно был применен для изучения изменений в развитии функциональной связи между областями мозга, которые составляют функциональные сети у детей старшего возраста и подростков. Эти исследования предлагают два типа связанных с возрастом изменений функциональной связи: снижение локальной связи между анатомически соседними, но функционально различными областями мозга по мере их интеграции в соответствующие сети, и увеличение дальней связи между узлами, которые составляют каждую сеть (Fair et al. и др., 2007; Келли и др., 2008). Они также подтверждают другие доказательства, предполагающие, что нейронные цепи, обслуживающие процессы внимания, созревают раньше, чем те, которые поддерживают социально-эмоциональное функционирование.
РАЗВИТИЕ НЕРВНЫХ ЦЕПЕЙ, ОБСЛУЖИВАЮЩИХ УПРАВЛЯЮЩИЕ ФУНКЦИИ
Обзор
По мере взросления дети и подростки ориентируются во все более сложной социальной среде со все большей автономией (Csikszentmihalyi et al, 1977). Чтобы преуспеть в этих новых жизненных задачах, они должны отточить и интегрировать многочисленные когнитивные и поведенческие процессы, каждый из которых, по-видимому, обслуживается определенной нейронной цепью. Эти когнитивные процессы более высокого порядка, в совокупности называемые «исполнительными функциями», включают в себя распределение внимания, рабочую память и саморегулируемый контроль. Хотя поведенческие проявления этих процессов широко изучены, мало что известно о развитии соответствующих им нейронных цепей, и еще меньше известно о том, как созревание мозга поддерживает координацию и интеграцию этих различных когнитивных областей. Нейровизуализационные исследования детей и подростков расширили нашу способность соотносить созревание сложных когнитивных процессов, составляющих исполнительные функции, с in vivo измерения структуры и функции мозга.
Развитие нейронных цепей для рабочей памяти
Рабочая память относится к способности поддерживать, обращать внимание и обновлять информацию, которая актуальна в данный момент и доступна «онлайн» для сознательной оценки и обработки (Baddele, 1986). В зрелом мозге рабочая память в значительной степени зависит от интактной дорсолатеральной префронтальной коры (ДЛПФК) (D’Esposito, 2007; Goldman-Rakic, 1994; Jonides et al, 1993; McCarthy et al, 19). 94; Оуэн и др., 1990; Петридес и др., 1993 г.; Уэйджер и Смит, 2003 г.). Рабочая память также задействует процессы внимания, которые базируются в латеральной и верхней лобно-теменной сети, включающей верхнюю лобную борозду, вентролатеральную префронтальную борозду (VLPFC), внутритеменную борозду и супрамаргинальную извилину (Champod and Petrides, 2007; Corbetta et al, 2008; Corbetta and Shulman). , 2002; Познер и Петерсен, 1990).
Элементарные возможности рабочей памяти наблюдаются у младенцев уже в возрасте 6 месяцев, но четкая способность к рабочей памяти развивается только в более позднем младенчестве (Brody, 1981; Алмаз, 1990; Миллар и Уотсон, 1979). В задаче Пиаже «А-не-В» (Piaget, 1954) младенец сначала наблюдает желаемый объект, например блестящую игрушку, помещенную в одну из двух лунок, которые обе закрыты. Затем младенцу разрешается открыть один колодец и, если он обнаружит желаемый объект, достать его. Затем испытание повторяется с объектом, спрятанным в том же колодце, и после нескольких успешных испытаний объект прячется во втором колодце. К 7–8 месяцам младенцы начинают успешно выполнять эту задачу, если им дают немедленно взять предмет, а не с задержкой всего в несколько секунд (Даймонд, 19 лет).90). К 9 месяцам младенцы могут выполнять эту задачу с задержкой. В последующие месяцы их способность хранить простую информацию в сети продолжает улучшаться (Diamond, 1990). Базовая способность к рабочей памяти прочно формируется к среднему детству (Tsujimoto et al, 2004). Рабочая память у детей легко перегружается при выполнении задач, требующих обработки информации в памяти или при наличии отвлекающих факторов (Davidson et al, 2006; Hitch, 2002), в то время как подростки способны выполнять более сложные задачи на рабочую память (Davidson et al, 2006). ; Деметриу и др., 2002 г.; Лусиана и Нельсон, 19 лет.98; Луна и др., 2004 г.; Шерф и др., 2006 г.; Суонсон, 1999).
Исследования фМРТ показывают, что при выполнении простых задач рабочей памяти на несколько классов информации у детей старшего возраста активируются лобно-теменные области (Casey et al, 1995; Crone et al, 2006; Durston et al, 2006; Geier et al, 2009). ; Клингберг и др., 2002; Конрад и др., 2005; Квон и др., 2002; Луна и др., 2001; Шерф и др., 2006; Томас и др., 1999). Таким образом, нейронные цепи для оперативной памяти уже установлены к этому возрасту. Тем не менее, дети старшего возраста не могут полностью и надлежащим образом задействовать лобно-теменную сеть во время выполнения более сложных задач рабочей памяти, таких как те, которые требуют манипулирования информацией в режиме онлайн (Crone et al, 2006; Konrad et al, 2005; Olesen et al, 2007). ) или поддерживать внимание на задаче, игнорируя отвлекающие факторы (Olesen et al, 2007). Вместо этого, по сравнению с подростками, дети задействуют вентромедиальные области, такие как хвостатое ядро и островок, во время выполнения более сложных задач рабочей памяти (Casey et al, 19).95; Кроун и др., 2006 г.; Дерстон и др., 2006 г.; Гейер и др., 2009 г.; Клингберг и др., 2002 г.; Конрад и др., 2005 г.; Квон и др., 2002 г.; Луна и др., 2001; Шерф и др., 2006 г.; Томас и др., 1999). Напротив, по сравнению со взрослыми подростки, по-видимому, задействуют пространственно более рассеянные участки лобной и теменной коры при задействовании процессов рабочей памяти (Durston et al, 2006; Konrad et al, 2005; Scherf et al, 2006). Таким образом, созревание нейронных цепей, поддерживающих процессы рабочей памяти, характеризуется более полным и последовательным задействованием лобно-теменных областей с увеличением сложности задач в период между детством и юностью, за которым следует пространственное уточнение этих областей коры между юностью и взрослой жизнью.
Развитие нейронных цепей, поддерживающих саморегуляцию
Дети часто должны выбирать между конкурирующими вариантами, такими как выполнение домашнего задания и игра с друзьями, или между вкусной закуской и сохранением места для ужина. Делая этот выбор, ребенок должен примирить конфликт между двумя конкурирующими вариантами, доступными в контексте ранее установленных ожиданий и правил, и, в конечном счете, должен подавлять автоматические импульсы к немедленному удовлетворению в пользу выбора, который менее автоматический, но который лучше. в соответствии с ценностями ребенка, семьи и общества. Таким образом, саморегуляция требует разработки и поддержания долгосрочных целей и правил, которые направляют и мотивируют поведение, а также выбора поведения, которое наилучшим образом согласовывает эти внутренние императивы с постоянно меняющимися внутренними и внешними непредвиденными обстоятельствами (Dubin et al, 2010). ).
В рамках широкой концепции саморегуляции когнитивный контроль относится к набору психических процессов, которые отвечают за выполнение, руководство и мониторинг желаемого поведения, подавляя при этом неуместные или невыгодные реакции (Dubin et al, 2010). Другие когнитивные процессы, такие как регуляция внимания, отслеживание конфликтов и ошибок, а также торможение реакции, служат когнитивному контролю (Bunge et al, 2002; Casey et al, 1997; Dubin et al, 2010; Rubia et al, 2000). Способность к когнитивному контролю или ее ранний прототип возникает в младенчестве. Например, в задаче Пиаже «А-не-В» многократное сокрытие объекта в первых двух лунках и разрешение его извлечения создают доминантную реакцию на поиск объекта в этой первой лунке (Piaget, 19).54). Способность правильно извлекать предмет после того, как он спрятан во второй лунке, которая появляется в возрасте 7–8 месяцев, частично зависит от торможения реакции или воздержания от доминантного поведения, связанного с проникновением в первую лунку (Даймонд, 1990; Пиаже, 1954).
Экспериментальные парадигмы, такие как задачи Stroop, Simon, Flanker, Go/No-Go и Stop-Signal, требуют подавления более автоматического поведения для выполнения менее автоматического. Регуляция внимания, торможение реакции, отслеживание конфликтов и ошибок — это когнитивные процессы, служащие когнитивному контролю и успешному выполнению задачи. Эффективность выполнения всех этих задач неуклонно улучшается на протяжении всего развития, но не приближается к уровню взрослых, по крайней мере, до позднего детства или ранней юности (Bunge et al, 2002; Casey et al, 19).97; Дэвидсон и др., 2006 г.; Луна и др., 2004 г.; Рубиа и др., 2000). Как и в случае с рабочей памятью, способность детей к саморегуляции может быть легко подавлена возрастающими требованиями к задачам.
У взрослых саморегуляция зависит от широких областей коры, таких как дополнительная моторная область, лобные поля глаза, передняя поясная кора, DLPFC, VLPFC/латеральная орбитофронтальная кора, а также височные и теменные области, все из которых связаны с полосатым телом в подкорке (Леунг и др. , 2000; Петерсон и др., 2002; Петерсон и др., 1999). Дети и взрослые задействуют эти лобно-полосатые цепи при выполнении задач, требующих саморегуляции (Adleman et al, 2002; Bunge et al, 2002; Casey et al, 1997; Luna et al, 2001; Peterson et al, 1999; Rubia et al. , 2000). Результаты исследований фМРТ позволяют предположить, что улучшение поведенческой способности к когнитивному контролю с возрастом связано с усилением активации лобных и полосатых цепей (рис. 7) (Adleman et al, 2002; Bunge et al, 2002; Casey et al, 1997; Luna). и др., 2001; Марш и др., 2006; Рубиа и др., 2000; Тамм и др., 2002). Имеющиеся данные свидетельствуют о том, что в мониторинге конфликтов участвует передняя поясная кора, в регуляции внимания участвуют ДЛПФК и теменная кора, подавление интерференции — ВЛПФК, а торможение реакции — теменные области. В задачах когнитивного контроля теменная активация продолжает усиливаться в подростковом возрасте, тогда как префронтальная активация продолжает усиливаться во взрослом возрасте (Adleman et al, 2002; Marsh et al, 2006; Rubia et al, 2006).
Разработка нейронных цепей для когнитивного контроля. Изображения представляют активацию мозга в поперечном сечении фМРТ развития задачи Струпа. ( а ) Воксельные корреляции возраста с активациями Струпа представлены в трансаксиальных срезах, расположенных сверху вниз (слева направо). (b) Групповые составные t-карты для изменения сигнала фМРТ, связанного с наименованием цветов в неконгруэнтных по сравнению с конгруэнтными стимулами для детей и взрослых. Увеличение сигнала во время неконгруэнтности по отношению к конгруэнтности кодируется желтым цветом, а уменьшение кодируется фиолетовым или синим цветом. Правое лобно-полосатое (ILPFC и Великий пост) увеличение активности, связанное с неконгруэнтными стимулами, постепенно проявлялось с возрастом. Таким образом, увеличение активности лобно-стриарных цепей с возрастом поддерживает улучшение когнитивного контроля в процессе развития у здоровых людей. PCC, задняя поясная кора; ACC, передняя поясная кора; VACC, вентральная передняя поясная кора; STG, верхняя височная извилина; Lnuc, чечевицеобразное ядро; LPFC, латеральная префронтальная кора; MPFC, мезиальная префронтальная кора; IFG, нижняя лобная извилина (Marsh et al, 2006) (Copyright 2006) John Wiley and Sons.
Слайд PowerPoint
Изображение в полный размер
Как саморегулируемая, так и система вознаграждения участвуют в разрешении конфликтов между внутренней гедонистической ценностью стимулов и их ценностью в более широкой структуре, которая включает интернализированные ценности и другие социальные влияния, личные цели и потенциальные риски среди другие последствия. У взрослых лобно-теменная и лобно-стриарная сети, включая дорсальный полосатый мозг, прилежащее ядро (NAcc), миндалевидное тело, OFC, латеральную и верхнюю области PFC и заднюю теменную кору, составляют нейронные цепи, обеспечивающие вознаграждение (Geier and Luna, 2009).; О’Доэрти, 2004). Исследования развития обработки вознаграждения с помощью фМРТ показали, что подростки отличаются от взрослых балансом между нисходящими механизмами внимания и когнитивного контроля, основанными главным образом на префронтальной коре, и восходящими мотивационными и эмоциональными реакциями на ситуации риска и вознаграждения, основанными на NAcc и миндалевидном теле. (Кейси и др., 2008 г.; Эрнст и Фадж, 2009 г.; Эрнст и др., 2005 г.; Гальван и др., 2006 г.; Спир, 2000 г.). Эти результаты согласуются с эмпирически подтвержденным мнением о том, что по сравнению со взрослыми подростки более мотивированы наградами, менее склонны к риску и легче поддаются влиянию сверстников (Arnett, 19).92; Стейнберг, 2007).
Возрастное увеличение активности лобно-теменных и лобно-полосатых цепей происходит одновременно с улучшением рабочей памяти и когнитивного контроля. Способность справляться со все более сложными задачами саморегуляции, особенно в условиях мотивационных или эмоциональных конфликтов, в значительной степени проявляется в подростковом возрасте, когда в лобно-теменных областях наблюдается истончение серого вещества коры (Gogtay et al, 2004; Sowell et al. , 2004). Точно так же улучшается выполнение задач исполнительного функционирования наряду с показателями созревания DTI в лобно-стриарных и лобно-теменных волокнистых путях (Barnea-Goraly et al, 2005; Olesen et al, 2003). Эти данные свидетельствуют о том, что изменения в структуре и функции лобно-теменных и лобно-стриарных цепей, связанные с развитием, могут быть связаны с ремоделированием синаптических связей и усилением связи между сетевыми узлами, что может обеспечить более активное участие и более эффективное функционирование этих цепей во все более сложных ситуациях. которые полагаются на исполнительную функцию.
Развитие нейронных цепей для исполнительных функций при психических расстройствах у детей
Психические заболевания, возникающие в детстве и подростковом возрасте, все чаще рассматриваются как нарушения в нейронных цепях, поддерживающие различные области когнитивного функционирования. Таким образом, полное понимание нормального развития мозговых цепей имеет решающее значение для выявления причин и улучшения лечения психических расстройств у детей и подростков. Например, обсессивно-компульсивное расстройство (ОКР), СДВГ и синдром Туретта (СТ) представляют собой педиатрические психические расстройства, которые часто возникают одновременно и характеризуются аномалиями в функционировании лобно-стриарных цепей, нарушающими саморегуляцию (Peterson et al, 2001). . Дети с ОКР, например, плохо справляются с задачами тормозного контроля, и этот дефицит коррелирует с клиническими симптомами (Maia et al, 2008). Анатомические и функциональные нейровизуализационные исследования показывают, что аномалии в орбитофронтальной коре, стриатуме и передней поясной коре нарушают функционирование лобно-стриарных цепей (Marsh et al, 2009).). Считается, что дефицит саморегуляции ведет к снижению способности направлять ресурсы внимания в сторону от навязчивых мыслей и подавлять принуждение к выполнению ритуалов (Chamberlain et al, 2005). Повышенная активация орбитофронтальной и передней поясной коры во время задач тормозного контроля может служить очагами дисфункции или может представлять собой компенсаторные реакции на подкорковые очаги дисфункции в лобно-стриарных цепях.
Обширные данные свидетельствуют о том, что у людей с СТ отсутствует способность активировать лобно-стриарные цепи и генерировать компенсаторную гиперплазию лобной коры, что в совокупности ухудшает их способность подавлять избыточную активность в сенсомоторных цепях. Эта сниженная ингибирующая способность затем ограничивает их способность подавлять тики (Marsh et al, 2009).). Напротив, у детей с СДВГ нарушен тормозной контроль, связанный с дорсальными лобно-стриарными цепями, а также нарушения процессов мотивации и вознаграждения, основанные на вентральных лобно-стриарных цепях, которые в совокупности могут вызывать дисфункцию обучения вознаграждению, толерантность к задержке и целенаправленную деятельность, которые характеризуют это расстройство (Sonuga-Barke, 2005; Sowell et al, 2003b). Следовательно, дифференциальные нарушения в лобно-стриарных цепях, по-видимому, по-разному опосредуют различные психические расстройства у детей и подростков.
ВЫВОДЫ И БУДУЩИЕ НАПРАВЛЕНИЯ
Развивающиеся экспериментальные подходы предоставили нам все более детальное представление о развитии цепей мозга как о длительной и даже пожизненной серии взаимосвязанных и частично перекрывающихся итерационных биологических процессов, которые следуют своим собственным временным направлениям. в разных областях мозга. Лежащие в основе клеточные процессы поддерживаются сложными молекулярными событиями, которые определяются сложными взаимодействиями генетических программ с воздействиями окружающей среды и опытом, которые также приводят к динамическому ремоделированию цепей на протяжении всей жизни. Правильное время и выполнение каждого из этих процессов критически важны для установления оптимальной функциональной способности нейронных цепей и, следовательно, могут также служить местом уязвимости для эффектов генетических аберраций и неблагоприятных воздействий окружающей среды.
Совокупность знаний, поддерживающих наше понимание нормального развития нейронных цепей у людей, тем не менее, все еще весьма несовершенна, поскольку она получена в основном из исследований поперечного сечения животных, в первую очередь грызунов. Эти отчеты дополняют относительно небольшое количество исследований на людях и приматах, в которых сравниваются показатели структуры и функций мозга в узких окнах развития. Таким образом, хотя исследования на грызунах и беспозвоночных предоставили бесценную информацию о генетических, молекулярных и клеточных процессах, которые направляют и регулируют развитие нейронных цепей, экстраполяция этих открытий на человека опасна и лишь частично устраняется скудными, но важными данными, полученными из исследований. нечеловекообразные приматы (Liu et al, 2008). Точно так же слишком частое использование данных о структуре и функциях нейронных цепей, полученных в один период развития, для вывода о предшествующих или последующих событиях развития имеет фундаментальные ограничения. Таким образом, наше понимание клеточных и молекулярных событий, которые способствуют созреванию нейронных цепей в детстве и подростковом возрасте у людей, остается предварительным и неполным.
Методы нейровизуализации позволяют проводить неинвазивные in vivo исследования структуры, состава и функций мозга, и эти методы внесли неоценимый вклад в наше понимание развития нейронных цепей. Хотя гистологические корреляты некоторых показателей визуализации были определены, микроскопические основы и биологическое значение многих других показателей еще предстоит определить. Кроме того, в большинстве педиатрических визуализирующих исследований на сегодняшний день делаются лонгитюдные выводы или выводы о развитии на основе поперечных данных, что может быть проблематично в лучшем случае по причинам, включающим систематическую ошибку, связанную с дифференцированной выборкой по возрастным группам, маскированием внутрииндивидуальных изменений путем индивидуальная изменчивость и трудности в определении сложных траекторий развития (Kraemer et al, 2000). Частично для решения этих трудностей с определением траекторий развития на основе данных поперечного сечения в последнее десятилетие появилось несколько продольных анатомических исследований развития мозга у детей и подростков. Эти исследования позволили непосредственно изучить траектории постнатального развития человеческого мозга и, предположительно, сделать более обоснованные выводы об их функциональных коррелятах.
Согласованное и интегрированное применение устоявшихся и новых экспериментальных методов может помочь устранить некоторые ограничения в современном состоянии знаний о развитии нейронных цепей. Мультимодальные визуализирующие исследования in vivo больших образцов здоровых образцов с последующим наблюдением в течение долгого времени облегчат не только прямое и беспристрастное изучение изменений в мозге, связанных с развитием, но и корреляцию биологической информации по методам визуализации, которая будет включать информацию о структуре мозга с использованием анатомической МРТ, связность аксонов с помощью DTI, функцию мозга и перфузию с помощью фМРТ, а также плотность нейронов и метаболизм с помощью MRS. Кроме того, применение современных клеточных и молекулярно-биологических методов для посмертного исследования мозга типично развивающихся детей позволит получить новое представление о гистологических и ультраструктурных особенностях развития нервной цепи. Сочетание этих исследований посмертной ткани с мультимодальными методами визуализации тех же образцов тканей позволит провести прямое исследование клеточных и молекулярных коррелятов данных посмертной визуализации, что значительно улучшит наше понимание клеточных основ и функциональных последствий изменений в развитии, выявленных в данные визуализации in vivo . Сочетание этих молекулярно-биологических методов с продольными исследованиями развития мозга у нечеловеческих приматов даст информацию, которая гораздо более актуальна, чем перекрестные исследования мозга грызунов, для улучшения нашего понимания развития человеческого мозга.
Изучение траекторий нормального развития мозговых цепей дает бесценную информацию о генезисе и биологических основах психопатологий, связанных с развитием. Выявление путей нормального развития мозга дает архимедову точку, с которой можно интерпретировать и понимать аберрантные пути развития мозга, вызывающие болезни. И наоборот, идентификация траекторий развития мозга, вызывающих болезни, дает нам информацию о нормальном развитии мозга, выявляя пути развития, которые наиболее чувствительны к нарушениям и, следовательно, имеют жизненно важное значение для построения здорового мозга. Вместе эти направления исследований должны улучшить наше понимание патогенеза психических заболеваний. Это улучшенное понимание должно, в свою очередь, помочь в планировании и разработке вмешательств, которые либо исправляют девиантные траектории развития, либо оптимизируют относительно нормальные траектории, чтобы гарантировать, что наши дети реализуют свой наибольший потенциал развития по мере взросления и взросления.
Ссылки
Адлеман Н.Е., Менон В., Блейси К.М., Уайт К.Д., Варсофски И.С., Гловер Г.Х. и др. (2002). Исследование развития с помощью фМРТ задачи Струпа на цветное слово. Нейроизображение 16 : 61–75.
ПабМед Google ученый
Allendoerfer KL, Shatz CJ (1994). Подпластинка, переходная неокортикальная структура: ее роль в развитии связей между таламусом и корой. Annu Rev Neurosci 17 : 185–218.
КАС пабмед Google ученый
Андерсен С.Л. (2003). Траектории развития мозга: точка уязвимости или окно возможностей? Neurosci Biobehav Rev 27 : 3–18.
ПабМед Google ученый
Арнетт Дж. (1992). Безрассудное поведение в подростковом возрасте: перспектива развития. Dev Rev 12 : 339–373.
Google ученый
Ashtari M, Cervellione KL, Hasan KM, Wu J, McIlree C, Kester H et al (2007). Развитие белого вещества в позднем подростковом возрасте у здоровых мужчин: исследование поперечного сечения диффузионного тензора. Нейроизображение 35 : 501–510.
ПабМед Google ученый
Аткинсон Дж. (1984). Зрительное развитие человека в первые 6 месяцев жизни. Обзор и гипотеза. Hum Neurobiol 3 : 61–74.
КАС пабмед Google ученый
Бэк С.А., Луо Н.Л., Боренштейн Н.С., Левин Дж.М., Вольпе Дж.Дж., Кинни Х.К. (2001). Поздние предшественники олигодендроцитов совпадают с периодом развития уязвимости для перинатального повреждения белого вещества человека. Дж Нейроски 21 : 1302–1312.
КАС пабмед ПабМед Центральный Google ученый
Бадделе АД (1986).
Рабочая память . Издательство Оксфордского университета: Нью-Йорк.Google ученый
Bagni C, Greenough WT (2005). От трафика мРНП до дисморфогенеза позвоночника: корни синдрома ломкой Х-хромосомы. Nat Rev Neurosci 6 : 376–387.
КАС пабмед Google ученый
Bansal R, Chung Y, Dong Z, Duan Y, Gerber A, Hao X et al (2007). Методы нейровизуализации в изучении психических расстройств детского возраста. В: Мартин А., Фолькмар Ф.Р., Льюис М. (ред.). Детская и подростковая психиатрия Льюиса: всеобъемлющий учебник , 4-е изд. Липпинкотт, Уильямс и Уилкинс: Балтимор, Мэриленд. стр. 214–234.
Google ученый
Баркович А.Дж., Кьос Б.О., Джексон Д.Э., Норман Д. (1988). Нормальное созревание головного мозга новорожденных и младенцев: МРТ при 1,5 Тл. Радиология 166 : 173–180.
КАС пабмед Google ученый
Барнеа-Горали Н., Менон В., Эккерт М., Тамм Л., Баммер Р., Карчемский А. и др. (2005). Развитие белого вещества в детстве и подростковом возрасте: исследование поперечного сечения диффузионного тензора. Кора головного мозга 15 : 1848–1854.
ПабМед Google ученый
Барцокис Г., Каммингс Дж.Л., Зультцер Д., Хендерсон В.В., Нухтерляйн К.Х., Минц Дж. (2003). Структурная целостность белого вещества у здоровых пожилых людей и пациентов с болезнью Альцгеймера: исследование магнитно-резонансной томографии. Arch Neurol 60 : 393–398.
ПабМед Google ученый
Bayer SA, Altman J (1990). Развитие слоя I и субпластины в неокортексе крысы. Exp Neurol 107 : 48–62.
КАС пабмед Google ученый
Беккер Л.
Э., Армстронг Д.Л., Чан Ф., Вуд М.М. (1984). Развитие дендритов в нейронах затылочной коры человека. Мозг Res 315 : 117–124.КАС пабмед Google ученый
Бекманн С., Делука М., Девлин Дж., Смит С. (2005). Исследования связности в состоянии покоя с использованием анализа независимых компонентов. Philos Trans R Soc B Biol Sci 360 : 1001–1013.
Google ученый
Белл М.А., Фокс Н.А. (1992). Отношения между электрической активностью лобного мозга и когнитивным развитием в младенчестве. Ребенок Дев 63 : 1142–1163.
КАС пабмед Google ученый
Бельмонте М.К., Аллен Г., Бекель-Митченер А., Буланже Л.М., Карпер Р.А., Уэбб С.Дж. (2004). Аутизм и аномальное развитие связей мозга. J Neurosci 24 : 9228–9231.
КАС пабмед ПабМед Центральный Google ученый
Бен Ари Ю.
, Хазипов Р., Лейнекугель Х., Кайяр О., Гайарса Д.Л. (1997). Рецепторы ГАМК А, NMDA и AMPA: регулируемое в процессе развития «управление втроем». Trends Neurosci 20 : 523–529.КАС пабмед Google ученый
Бен Башат Д., Бен Сира Л., Грайф М., Пианка П., Хендлер Т., Коэн И. и др. (2005). Нормальное развитие белого вещества от младенчества до взрослой жизни: сравнение тензора диффузии и диффузионно-взвешенных МРТ-изображений с высоким значением b. J Magn Reson Imaging 21 : 503–511.
ПабМед Google ученый
Бенеш Ф.М., Черепаха М., Хан Ю., Фарол П. (1994). Миелинизация ключевой зоны реле в формировании гиппокампа происходит в мозге человека в детстве, подростковом и взрослом возрасте. Arch General Psychiatry 51 : 477–484.
КАС пабмед Google ученый
Берарди Н.
, Пиццоруссо Т., Маффеи Л. (2000). Критические периоды сенсорного развития. Curr Opin Neurobiol 10 : 138–145.КАС пабмед Google ученый
Berman JI, Mukherjee P, Partridge SC, Miller SP, Ferriero DM, Barkovich AJ et al (2005). Количественная диффузионно-тензорная МРТ-волоконная трактография сенсомоторного развития белого вещества у недоношенных детей. Нейроизображение 27 : 862–871.
ПабМед Google ученый
Bhardwaj RD, Curtis MA, Spalding KL, Buchholz BA, Fink D, Bjork-Eriksson T et al (2006). Неокортикальный нейрогенез у людей ограничен развитием. Proc Natl Acad Sci USA 103 : 12564–12568.
КАС пабмед ПабМед Центральный Google ученый
Биндер Дж. Р., Фрост Дж. А., Хаммеке Т. А., Беллгован П. С., Рао С. М.
, Кокс Р. В. (1999). Концептуальная обработка во время сознательного состояния покоя. Функциональное МРТ исследование. J Cogn Neurosci 11 : 80–95.КАС пабмед Google ученый
Бинцеггер Т., Дуглас Р., Мартин К.А. (2004). Количественная карта цепи первичной зрительной коры кошки. J Neurosci 24 : 8441–8453.
КАС пабмед ПабМед Центральный Google ученый
Бисвал Б., Йеткин Ф.З., Хотон В.М., Хайд Дж.С. (1995). Функциональная связь в моторной коре покоящегося мозга человека с помощью эхо-планарной МРТ. Magn Reson Med 34 : 537–541.
КАС пабмед Google ученый
Блашке А.Дж., Стейли К., Чун Дж. (1996). Широко распространенная запрограммированная гибель клеток в пролиферативных и постмитотических областях коры головного мозга плода.
Разработка 122 : 1165–1174.КАС пабмед Google ученый
Буржуазный JP (1997). Синаптогенез, гетерохрония и эпигенез в неокортексе млекопитающих. Acta Paediatr Suppl 422 : 27–33.
КАС пабмед Google ученый
Буржуа Дж. П., Гольдман-Ракич П. С., Ракич П. (1994). Синаптогенез в префронтальной коре макак-резусов. Кора головного мозга 4 : 78–96.
КАС пабмед Google ученый
Буржуа Дж. П., Ракич П. (1993). Изменения синаптической плотности в первичной зрительной коре макаки от эмбриональной до взрослой стадии. J Neurosci 13 : 2801–2820.
КАС пабмед ПабМед Центральный Google ученый
Броуди Б.А., Кинни Х.К., Кломан А.С., Жиль Ф.Х. (1987).
Последовательность миелинизации центральной нервной системы в младенчестве человека. I. Вскрытие миелинизации. J Neuropathol Exp Neurol 46 : 283–301.КАС пабмед Google ученый
Броды Л.Р. (1981). Кратковременная зрительная память в младенчестве. Ребенок Дев 52 : 242–250.
КАС пабмед Google ученый
Бунге С.А., Дудукович Н.М., Томасон М.Е., Вайдья С.Дж., Габриэли Д.Д. (2002). Вклад незрелых лобных долей в когнитивный контроль у детей: данные фМРТ. Нейрон 33 : 301–311.
КАС пабмед ПабМед Центральный Google ученый
Быстрон И., Блейкмор С., Ракич П. (2008). Развитие коры головного мозга человека: новый взгляд на комитет Боулдера. Nat Rev Neurosci 9 : 110–122.
КАС пабмед Google ученый
Кэди Э.
Б., Пенрис Дж., Амесс П.Н., Лорек А., Вилезинска М., Олдридж РФ и др. (1996). Концентрации лактата, N-ацетиласпартата, холина и креатина и спин-спиновая релаксация в таламической и затылочно-теменной областях развивающегося головного мозга человека. Magn Reson Med 36 : 878–886.КАС пабмед Google ученый
Карпентер Г.Л., Stacks AM (2009). Влияние на развитие воздействия насилия со стороны интимного партнера в раннем детстве: обзор литературы. Children Youth Serv Rev 31 : 831–839.
Google ученый
Кейси Б.Дж., Коэн Дж.Д., Джеззард П., Тернер Р., Нолл Д.К., Трейнор Р.Дж. и др. (1995). Активация префронтальной коры у детей во время задания на непространственную рабочую память с помощью функциональной МРТ. Нейроизображение 2 : 221–229.
КАС пабмед Google ученый
Кейси Б.
Дж., Джонс Р.М., Заяц Т.А. (2008). Подростковый мозг. Ann NY Acad Sci 1124 : 111–126.КАС пабмед Google ученый
Casey BJ, Trainor RJ, Orendi JL, Schubert AB, Nystrom LE, Giedd JN et al (1997). Эволюционное функциональное МРТ-исследование префронтальной активации во время выполнения задачи Go-No-Go. J Cogn Neurosci 9 : 835–847.
КАС пабмед Google ученый
Caspi A, Sugden K, Moffitt TE, Taylor A, Craig IW, Harrington H et al (2003). Влияние жизненного стресса на депрессию: модерация полиморфизмом в гене 5-НТТ. Наука 301 : 386–389.
КАС пабмед Google ученый
Чемберлен С.Р., Блэквелл А.Д., Файнберг Н.А., Роббинс Т.В., Саакян Б.Дж. (2005). Нейропсихология обсессивно-компульсивного расстройства: важность сбоев в когнитивном и поведенческом торможении в качестве возможных эндофенотипических маркеров.
Neurosci Biobehav Rev 29 : 399–419.КАС пабмед Google ученый
Чампод А.С., Петридес М. (2007). Диссоциируемые роли задней теменной и префронтальной коры в процессах манипулирования и мониторинга. Proc Natl Acad Sci USA 104 : 14837–14842.
КАС пабмед ПабМед Центральный Google ученый
Changeux JP, Danchin A (1976). Избирательная стабилизация развивающихся синапсов как механизм спецификации нейронных сетей. Природа 264 : 705–712.
КАС пабмед Google ученый
Чао Д., Ма Л., Шен К. (2009). Временные межклеточные взаимодействия при формировании нейронной цепи. Nat Rev Neurosci 10 : 262–271.
КАС пабмед ПабМед Центральный Google ученый
Чугани ХТ (1994).
Развитие регионального метаболизма глюкозы в головном мозге в связи с поведением и пластичностью. В: Доусон Г., Фишер К.В. (ред.). Человеческое поведение и развивающийся мозг . Гилфорд: Нью-Йорк. стр. 153–175.Google ученый
Чугани ХТ, Фелпс МЭ (1986). Возрастные изменения функции головного мозга у детей раннего возраста, определяемые с помощью позитронно-эмиссионной томографии с 18ФДГ. Наука 231 : 840–843.
КАС пабмед Google ученый
Чугани Х.Т., Фелпс М.Е., Мацциотта Дж.К. (1987). Исследование функционального развития головного мозга человека с помощью позитронно-эмиссионной томографии. Энн Нейрол 22 : 487–497.
КАС пабмед Google ученый
Цитри А, Маленко Р (2008). Синаптическая пластичность: множественные формы, функции и механизмы. Нейропсихофармакология 33 : 18–41.
ПабМед Google ученый
Корбетта М., Патель Г., Шульман Г.Л. (2008). Система переориентации человеческого мозга: от среды к теории сознания. Нейрон 58 : 306–324.
КАС пабмед ПабМед Центральный Google ученый
Корбетта М., Шульман Г.Л. (2002). Контроль целенаправленного и стимулированного внимания в головном мозге. Nat Rev Neurosci 3 : 201–215.
КАС пабмед Google ученый
Courchesne E, Chisum HJ, Townsend J, Cowles A, Covington J, Egaas B et al (2000). Нормальное развитие мозга и старение: количественный анализ на in vivo МРТ здоровых добровольцев. Радиология 216 : 672–682.
КАС пабмед Google ученый
Courchesne E, Karns CM, Davis HR, Ziccardi R, Carper RA, Tigue ZD et al (2001).
Необычные модели роста мозга в раннем возрасте у пациентов с аутистическим расстройством: исследование МРТ. Неврология 57 : 245–254.КАС пабмед Google ученый
Коуэн В.М., Фосетт Дж.В., О’Лири Д.Д., Стэнфилд Б.Б. (1984). Регрессивные события в нейрогенезе. Наука 225 : 1258–1265.
КАС пабмед Google ученый
Кроун Э., Венделкен С., Донохью С., ван Лейенхорст Л., Бунге С.А. (2006). Нейрокогнитивное развитие способности манипулировать информацией в рабочей памяти. Proc Natl Acad Sci USA 103 : 9315–9320.
КАС пабмед ПабМед Центральный Google ученый
Чиксентмихайи М., Ларсон Р., Прескотт С. (1977). Экология подростковой деятельности и переживаний. Молодежь подростков 6 : 281–294.
КАС Google ученый
Д’Эспозито М (2007).
От когнитивных к нейронным моделям рабочей памяти. Philos Trans R Soc Lond B Biol Sci 362 : 761–772.ПабМед ПабМед Центральный Google ученый
Дамуазо Дж., Ромбоутс С., Баркхоф Ф., Шелтенс П., Стэм С., Смит С. и др. (2006). Согласованные сети состояния покоя у здоровых субъектов. Proc Natl Acad Sci 103 : 13848–13853.
КАС пабмед ПабМед Центральный Google ученый
Дэвидсон М.С., Амсо Д., Андерсон Л.С., Даймонд А. (2006). Развитие когнитивного контроля и исполнительных функций от 4 до 13 лет: данные манипуляций с памятью, торможения и переключения задач. Нейропсихология 44 : 2037–2078.
ПабМед ПабМед Центральный Google ученый
де Грааф-Питерс В.Б., Хаддерс-Альгра М. (2006). Онтогенез центральной нервной системы человека: что и когда происходит? Ранний Хум Дев 82 : 257–266.
ПабМед Google ученый
де ла Роса Э.Дж., де Пабло Ф. (2000). Гибель клеток в раннем развитии нервной системы: за пределами нейротрофической теории. Trends Neurosci 23 : 454–458.
КАС пабмед Google ученый
de Vries JI, Visser GH, Prechtl HF (1985). Возникновение поведения плода. II. Количественные аспекты. Ранний Хум Дев 12 : 99–120.
КАС пабмед Google ученый
ДеФелипе Дж., Фаринас И. (1992). Пирамидный нейрон коры головного мозга: морфологическая и химическая характеристика синаптических входов. Prog Neurobiol 39 : 563–607.
КАС пабмед Google ученый
Декабан АС (1978). Изменения массы мозга в течение жизни человека: отношение массы мозга к росту и массе тела.
Энн Нейрол 4 : 345–356.КАС пабмед Google ученый
Делука М., Бекманн С., Дестефано Н., Мэтьюз П., Смит С. (2006). Сети состояния покоя фМРТ определяют различные режимы взаимодействия на расстоянии в человеческом мозгу. НейроИзображение 29 : 1359–1367.
КАС Google ученый
Деметриу А., Христу С., Спанудис Г., Платсиду М. (2002). Развитие мыслительной обработки: работоспособность, рабочая память и мышление. Monogr Soc Res Child Dev 67 : 1–156.
Google ученый
Девис Д., Бьянкалана В., Руссо Ф., Буэ Дж., Мандель Дж.Л., Оберле И. (1992). Анализ полных ломких Х-мутаций в тканях плода и монозиготных близнецов указывает на то, что аномальное метилирование и соматическая гетерогенность устанавливаются на ранних этапах развития. Am J Med Genet 43 : 208–216.
КАС пабмед Google ученый
Алмаз А (1990). Развитие и нейронные основы функций памяти, индексированные AB и задачами с отсроченным ответом, у младенцев человека и детенышей обезьян. Ann NY Acad Sci 608 : 267–309; обсуждение 309–317.
КАС пабмед Google ученый
Алмаз А (1995). Руководство по изучению взаимосвязей поведения мозга в процессе развития. В: Харви Л., Айзенберг Х. (ред.). Дисфункция лобных долей . Издательство Оксфордского университета: Нью-Йорк. стр. 339–378.
Google ученый
Дуглас Р.Дж., Мартин К.А. (2004). Нейронные цепи неокортекса. Annu Rev Neurosci 27 : 419–451.
КАС пабмед Google ученый
Дубин М.Дж., Майя ТВ, Петерсон Б.С. (2010). Когнитивный контроль на службе саморегуляции.
В: Koob GF, Le Moal M, Thompson RF (eds). Энциклопедия поведенческой неврологии . Эльзевир: Амстердам, Нидерланды.Google ученый
Дерстон С., Дэвидсон М.С., Тоттенхэм Н., Гальван А., Спайсер Д., Фосселла Д.А. и др. (2006). Переход от диффузной к очаговой активности коры по мере развития. Dev Sci 9 : 1–8.
ПабМед Google ученый
Эрлих И., Малинов Р. (2004). Постсинаптическая плотность 95 контролирует включение AMPA-рецепторов во время долговременной потенциации и управляемой опытом синаптической пластичности. J Neurosci 24 : 916–927.
КАС пабмед ПабМед Центральный Google ученый
Эрнст М., Фадж Дж. (2009). Нейробиологическая модель развития мотивированного поведения: анатомия, связь и онтогенез триадных узлов. Neurosci Biobehav Rev 33 : 367–382.
ПабМед Google ученый
Ernst M, Nelson E, Jazbec S, McClure E, Monk C, Leibenluft E et al (2005). Миндалевидное тело и прилежащее ядро в ответ на получение и пропуск усиления у взрослых и подростков. НейроИзображение 25 : 1279–1291.
ПабМед Google ученый
Эйр Дж. А., Тейлор Дж. П., Виллагра Ф., Смит М., Миллер С. (2001). Доказательства зависимости от активности корково-спинномозговых проекций во время развития человека. Неврология 57 : 1543–1554.
КАС пабмед Google ученый
Fair D, Dosenbach N, Church JA, Cohen A, Brahmbhatt S, Miezin F и др. (2007). Развитие отдельных сетей управления посредством разделения и интеграции. Proc Natl Acad Sci 104 : 13507–13512.
КАС пабмед ПабМед Центральный Google ученый
Файнберг И.
(1982). Шизофрения: вызвана нарушением запрограммированной синаптической элиминации в подростковом возрасте? J Psychiatr Res 17 : 319–334.ПабМед Google ученый
Финлей Б.Л., Дарлингтон Р.Б. (1995). Связанные закономерности в развитии и эволюции мозга млекопитающих. Наука 268 : 1578–1584.
КАС пабмед Google ученый
Fogliarini C, Chaumoitre K, Chapon F, Fernandez C, Levrier O, Figarella-Branger D et al (2005). Оценка созревания коры с помощью пренатальной МРТ. Часть I: нормальное созревание коры. Евро Радиол 15 : 1671–1685.
ПабМед Google ученый
Фокс М., Корбетта М., Снайдер А.З., Винсент Дж.Л., Райхл М. (2006). Спонтанная активность нейронов отличает дорсальную и вентральную системы внимания человека. Proc Natl Acad Sci USA 103 : 10046–10051.
КАС пабмед ПабМед Центральный Google ученый
Фокс М., Райхл М. (2007). Спонтанные колебания активности головного мозга наблюдаются при функциональной магнитно-резонансной томографии. Nat Rev Neurosci 8 : 700–711.
КАС пабмед Google ученый
Фокс М., Снайдер А.З., Винсент Дж.Л., Корбетта М., Ван Эссен Д.К., Райхл М. (2005). Человеческий мозг внутренне организован в виде динамических, антикоррелированных функциональных сетей. Proc Natl Acad Sci USA 102 : 9673–9678.
КАС пабмед ПабМед Центральный Google ученый
Фрэнсис Д., Диорио Дж., Лю Д., Мини М.Дж. (1999). Негеномная передача через поколения материнского поведения и реакции на стресс у крыс. Наука 286 : 1155–1158.
КАС пабмед Google ученый
Франссон П.
, Скиольд Б., Хорш С., Норделл А., Бленноу М., Лагеркранц Х. и др. (2007). Сети состояния покоя в мозге младенцев. Proc Natl Acad Sci USA 104 : 15531–15536.КАС пабмед ПабМед Центральный Google ученый
Friauf E, McConnell SK, Shatz CJ (1990). Функциональные синаптические цепи в субпластине во время внутриутробного и раннего постнатального развития зрительной коры кошки. J Neurosci 10 : 2601–2613.
КАС пабмед ПабМед Центральный Google ученый
Galvan A, Hare TA, Parra CE, Penn J, Voss H, Glover G и др. (2006). Более раннее развитие прилежащей коры относительно орбитофронтальной коры может лежать в основе рискованного поведения у подростков. J Neurosci 26 : 6885–6892.
КАС пабмед ПабМед Центральный Google ученый
Gao W, Lin W, Chen Y, Gerig G, Smith JK, Jewells V et al (2009).
Временное и пространственное развитие созревания аксонов и миелинизации белого вещества в развивающемся мозге. AJNR Am J Нейрорадиол 30 : 290–296.КАС пабмед ПабМед Центральный Google ученый
Гарель С., Шантрел Э., Брис Х., Эльмалех М., Лутон Д., Ури Дж. Ф. и др. (2001). Кора головного мозга плода: нормальные гестационные ориентиры, выявленные с помощью пренатальной МРТ. AJNR Am J Нейрорадиол 22 : 184–189.
КАС пабмед ПабМед Центральный Google ученый
Гейер С., Луна Б. (2009). Созревание обработки стимулов и когнитивного контроля. Pharmacol Biochem Behav 89 : 1–10.
Google ученый
Гейер К.Ф., Гарвер К., Тервиллигер Р., Луна Б. (2009). Развитие поддержания рабочей памяти. J Нейрофизиол 101 : 84–99.
ПабМед Google ученый
Гашгаи Х.Т., Лай С., Антон Э.С. (2007). Миграция нейронов во взрослом мозге: мы уже там? Nat Rev Neurosci 8 : 141–151.
КАС пабмед Google ученый
Гош А., Антонини А., МакКоннелл С.К., Шац С.Дж. (1990). Потребность нейронов подпластины в формировании таламокортикальных связей. Природа 347 : 179–181.
КАС пабмед Google ученый
Гош А., Шац С.Дж. (1992). Участие нейронов субпластины в формировании колонок окулярного доминирования. Наука 255 : 1441–1443.
КАС пабмед Google ученый
Гидд Дж. (1999). Развитие мозга, IX: рост человеческого мозга. Am J Психиатрия 156 : 4.
CAS пабмед Google ученый
Giedd J, Blumenthal J, Jeffries NO, Castellanos FX, Liu H, Zijdenbos A et al (1999).
Развитие мозга в детстве и подростковом возрасте: продольное МРТ-исследование. Nat Neurosci 2 : 861–863.КАС пабмед Google ученый
Giedd JN, Snell JW, Lange N, Rajapakse JC, Casey BJ, Kozuch PL et al (1996a). Количественная магнитно-резонансная томография развития человеческого мозга: возраст от 4 до 18 лет. Кора головного мозга 6 : 551–560.
КАС пабмед Google ученый
Giedd JN, Vaituzis AC, Hamburger SD, Lange N, Rajapakse JC, Kaysen D et al (1996b). Количественная МРТ височной доли, миндалевидного тела и гиппокампа в нормальном развитии человека: возраст 4–18 лет. J Comp Neurol 366 : 223–230.
КАС пабмед Google ученый
Гилберт CD (1983). Микросхема зрительной коры. Annu Rev Neurosci 6 : 217–247.
КАС пабмед Google ученый
Gilmore JH, Lin W, Corouge I, Vetsa YS, Smith JK, Kang C et al (2007a). Раннее постнатальное развитие мозолистого тела и кортикоспинального белого вещества оценивают с помощью количественной трактографии. AJNR Am J Нейрорадиол 28 : 1789–1795.
КАС пабмед ПабМед Центральный Google ученый
Гилмор Дж. Х., Лин В., Гериг Г. (2006). Развитие мозга плода и новорожденного. Am J Психиатрия 163 : 2046.
PubMed Google ученый
Gilmore JH, Lin W, Prastawa MW, Looney CB, Vetsa YS, Knickmeyer RC et al (2007b). Региональный рост серого вещества, половой диморфизм и церебральная асимметрия в неонатальном мозге. J Neurosci 27 : 1255–1260.
КАС пабмед ПабМед Центральный Google ученый
Джорджио А.
, Уоткинс К.Е., Дуо Г., Джеймс А.С., Джеймс С., Де Стефано Н. и др. (2008). Изменения микроструктуры белого вещества в подростковом возрасте. НейроИзображение 39 : 52–61.КАС пабмед Google ученый
Жирар Н., Рейбо К., дю Лак П. (1991). МРТ-исследование миелинизации головного мозга. J Нейрорадиол 18 : 291–307.
КАС пабмед Google ученый
Года Ю., Дэвис Г.В. (2003). Механизмы сборки и разборки синапсов. Нейрон 40 : 243–264.
КАС пабмед Google ученый
Гогтай Н., Гидд Дж., Луск Л., Хаяши К.М., Гринштейн Д., Вайтузис А.С. и др. (2004). Динамическое картирование развития коры головного мозга человека в период от детства до раннего взросления. Proc Natl Acad Sci USA 101 : 8174–8179.
КАС пабмед ПабМед Центральный Google ученый
Гольке Дж.
, Гриффит В., Фаустман Э. (2006). Вычислительные модели нейрогенеза неокортекса и запрограммированной гибели клеток у развивающихся мышей, обезьян и человека. Кора головного мозга 17 : 2433–2442.Google ученый
Goldman-Rakic PS (1994). Проблема памяти в изучении префронтальных функций. В: Тьерри AM (ред.). Двигательные и когнитивные функции префронтальной коры . Springer-Verlag: Берлин. стр. 112–122.
Google ученый
Гомес М.Р., Сэмпсон Дж.Р., Уиттемор В.Х. (1999). Туберозный склероз , 3-е изд. Издательство Оксфордского университета: Нью-Йорк.
Google ученый
Гудкин Ф. (1980). Развитие зрелых паттернов координации головы и глаз у младенцев. Ранний Хум Дев 4 : 373–386.
КАС пабмед Google ученый
Грациани Л.
Дж., Вайцман Э.Д., Веласко М.С. (1968). Неврологическое созревание и слуховые вызванные реакции у детей с низкой массой тела при рождении. Педиатрия 41 : 483–494.КАС пабмед Google ученый
Грейциус М.Д., Краснов Б., Рейсс А.Л., Менон В. (2003). Функциональная связь в мозге в состоянии покоя: сетевой анализ гипотезы режима по умолчанию. Proc Natl Acad Sci USA 100 : 253–258.
КАС пабмед Google ученый
Грессенс П. (2000). Механизмы и нарушения миграции нейронов. Pediatr Res 48 : 725–730.
КАС пабмед Google ученый
Гупта Р.К., Хасан К.М., Триведи Р., Прадхан М., Дас В., Парих Н.А. и др. (2005). Диффузионно-тензорная визуализация развивающегося головного мозга человека. J Neurosci Res 81 : 172–178.
КАС пабмед Google ученый
Гуснард Д.А., Акбудак Э., Шульман Г.Л., Райхле М.Е. (2001). Медиальная префронтальная кора и самореферентная умственная деятельность: отношение к режиму работы мозга по умолчанию. Proc Natl Acad Sci USA 98 : 4259–4264.
КАС пабмед ПабМед Центральный Google ученый
Хэмпсон М., Петерсон Б.С., Скудларски П., Гейтенби Дж. К., Гор Дж. К. (2002). Обнаружение функциональной связи с использованием временных корреляций на МР-изображениях. Hum Brain Map 15 : 247–262.
ПабМед ПабМед Центральный Google ученый
Ханаока С., Такашима С., Морока К. (1998). Исследование созревания головного мозга ребенка с помощью 31P-MRS. Pediatr Neurol 18 : 305–310.
КАС пабмед Google ученый
Хардан А.
Ю., Муддасани С., Вемулапалли М., Кешаван М.С., Миншью Н.Дж. (2006). МРТ-исследование увеличения толщины коры при аутизме. Am J Психиатрия 163 : 1290–1292.ПабМед ПабМед Центральный Google ученый
Хаттен М.Э. (1993). Роль миграции в развитии нейронов центральной нервной системы. Curr Opin Neurobiol 3 : 38–44.
КАС пабмед Google ученый
Хаттен М.Э. (1999). Миграция нейронов центральной нервной системы. Annu Rev Neurosci 22 : 511–539.
КАС пабмед Google ученый
Hazlett HC, Poe M, Gerig G, Smith RG, Provenzale J, Ross A et al (2005). Магнитно-резонансная томография и исследование размера мозга по окружности головы при аутизме: от рождения до 2 лет. Arch General Psychiatry 62 : 1366–1376.
ПабМед Google ученый
Hermoye L, Saint-Martin C, Cosnard G, Lee SK, Kim J, Nassogne MC et al (2006).
Педиатрическая диффузионная тензорная визуализация: нормальная база данных и наблюдение за созреванием белого вещества в раннем детстве. Нейроизображение 29 : 493–504.ПабМед Google ученый
Сцепка GJ (2002 г.). Эволюционные изменения рабочей памяти: многокомпонентный взгляд. В: Граф П., Охта Н. (ред.). Продолжительность жизни Развитие обработки человека . MIT Press: Кембридж, Массачусетс, стр. 15–37.
Google ученый
Холден К.Р. (2007 г.). Недоедание и развитие мозга. Семин Клин Нейрол 6 : 19–36.
Google ученый
Крючки BM, Chen C (2006). Различная роль спонтанной и зрительной активности в ремоделировании ретиногенетического синапса. Нейрон 52 : 281–291.
КАС пабмед Google ученый
Хорска А.
, Кауфманн В.Е., Брант Л.Дж., Найду С., Харрис Дж.К., Баркер П.Б. (2002). In vivo количественное протонное MRSI исследование развития мозга с детства до подросткового возраста. J Magn Reson Imaging 15 : 137–143.ПабМед Google ученый
Huang H, Zhang J, Wakana S, Zhang W, Ren T, Richards L et al (2006). Развитие белого и серого вещества головного мозга плода, новорожденного и ребенка. НейроИзображение 33 : 27–38.
ПабМед Google ученый
Хьюбел Д. Х., Визель Т. Н. (1962). Рецептивные поля, бинокулярное взаимодействие и функциональная архитектура зрительной коры кошек. J Physiol 160 : 106–154.
КАС пабмед ПабМед Центральный Google ученый
Хубер К.М., Галлахер С.М., Уоррен С.Т., Медведь М.Ф. (2002). Изменение синаптической пластичности в мышиной модели ломкой X умственной отсталости.
Proc Natl Acad Sci USA 99 : 7746–7750.КАС пабмед ПабМед Центральный Google ученый
Губерман А.Д. (2007 г.). Механизмы развития глазоспецифических зрительных цепей. Curr Opin Neurobiol 17 : 73–80.
КАС пабмед Google ученый
Хьюисман Т.А., Мартин Э., Кубик-Хуч Р., Маринчек Б. (2002). Магнитно-резонансная томография головного мозга плода: технические аспекты и нормальное развитие мозга. Евро Радиол 12 : 1941–1951.
ПабМед Google ученый
Hüppi PS, Warfield S, Kikinis R, Barnes PD, Zientara GP, Jolesz FA et al (1998). Количественная магнитно-резонансная томография развития головного мозга у недоношенных и доношенных новорожденных. Энн Нейрол 43 : 224–235.
ПабМед Google ученый
Huttenlocher PR (1979).
Синаптическая плотность в лобной коре человека — изменения в развитии и последствия старения. Мозг Res 163 : 195–205.КАС пабмед Google ученый
Huttenlocher PR (1984). Устранение синапсов и пластичность в развивающейся коре головного мозга человека. Am J Ment Defic 88 : 488–496.
КАС пабмед Google ученый
Huttenlocher PR (1990). Морфометрическое исследование развития коры головного мозга человека. Нейропсихология 28 : 517–527.
КАС пабмед Google ученый
Huttenlocher PR, Dabholkar AS (1997). Региональные различия в синаптогенезе в коре головного мозга человека. J Comp Neurol 387 : 167–178.
КАС пабмед Google ученый
Huttenlocher PR, De Courten C, Garey LJ, Van der Loos H (1982a).
Развитие синапсов в коре головного мозга человека. Int J Neurol 16–17 : 144–154.ПабМед Google ученый
Huttenlocher PR, de Courten C, Garey LJ, Van der Loos H (1982b). Синаптогенез в зрительной коре человека — свидетельство элиминации синапсов во время нормального развития. Neurosci Lett 33 : 247–252.
КАС пабмед Google ученый
Иай М., Ямамура Т., Такашима С. (1997). Ранняя экспрессия протеолипидного белка в головном мозге плода и младенца человека. Pediatr Neurol 17 : 235–239.
КАС пабмед Google ученый
Инноченти GM (1981). Рост и изменение формы аксонов при установлении зрительных мозолистых связей. Наука 212 : 824–827.
КАС пабмед Google ученый
Исаак Дж.
Т.Р., Крейр М.С., Николл Р.А., Маленко Р.С. (1997). Молчащие синапсы во время развития таламокортикального входа. Нейрон 18 : 269–280.КАС пабмед Google ученый
Джерниган Т.Л., Траунер Д.А., Хесселинк Дж.Р., Таллал П.А. (1991). Наблюдается созревание головного мозга человека in vivo в подростковом возрасте. Мозг 114 (Часть 5): 2037–2049.
ПабМед Google ученый
Jessell TM, Sanes JR (2000). Разработка. Десятилетие развивающегося мозга. Curr Opin Neurobiol 10 : 599–611.
КАС пабмед Google ученый
Джонидес Дж., Смит Э.Э., Кёппе Р.А., Аух Э., Миношима С., Минтун М.А. (1993). Пространственная рабочая память у людей по данным ПЭТ. Природа 363 : 623–625.
КАС пабмед Google ученый
Жуэ М.
, Розенталь А., Армстронг Г., Макфарлейн Дж., Стивенсон Р., Патерсон Дж. и др. (1994). Х-сцепленная спастическая параплегия (SPG1), синдром MASA и Х-сцепленная гидроцефалия являются результатом мутаций в гене L1. Нат Жене 7 : 402–407.КАС пабмед Google ученый
Кандель Э.Р., Шварц Дж.Х., Санес Дж.Р. (2000). Сенсорный опыт и тонкая настройка синаптических связей. В: Kandel ER, Schwartz JH, Jessell TM (ред.). Принципы нейробиологии , 4-е изд. Appleton & Lange: Стэмфорд, Коннектикут, стр. 1115–1130.
Google ученый
Канольд ПО (2004). Переходные микросхемы, образованные нейронами субпластин, и их роль в функциональном развитии таламокортикальных связей. Нейроотчет 15 : 2149–2153.
ПабМед Google ученый
Кац PS (2007). Эволюция и развитие нейронных цепей у беспозвоночных.
Curr Opin Neurobiol 17 : 59–64.КАС пабмед Google ученый
Келли А., Ди Мартино А., Уддин Л., Шехзад З., Джи Д., Рейсс П. и др. (2008). Развитие функциональной связности передней поясной извилины с позднего детства до раннего взросления. Кора головного мозга 19 : 640–657.
ПабМед Google ученый
Кинни Х.К., Броди Б.А., Кломан А.С., Жиль Ф.Х. (1988). Последовательность миелинизации центральной нервной системы в младенчестве человека. II. Паттерны миелинизации у вскрытых младенцев. J Neuropathol Exp Neurol 47 : 217–234.
КАС пабмед Google ученый
Клингберг Т., Форссберг Х., Вестерберг Х. (2002). Повышенная мозговая активность в лобной и теменной коре лежит в основе развития зрительно-пространственной рабочей памяти в детстве.
J Cogn Neurosci 14 : 1–10.ПабМед Google ученый
Клингберг Т., Вайдья С.Дж., Габриэли Д.Д., Мозли М.Е., Хедехус М. (1999). Миелинизация и организация лобного белого вещества у детей: диффузионно-тензорное МРТ исследование. Нейроотчет 10 : 2817–2821.
КАС пабмед Google ученый
Knickmeyer RC, Gouttard S, Kang C, Evans D, Wilber K, Smith JK et al (2008). Структурное МРТ-исследование развития головного мозга человека от рождения до 2 лет. J Neurosci 28 : 12176–12182.
КАС пабмед ПабМед Центральный Google ученый
Konrad K, Neufang S, Thiel CM, Specht K, Hanisch C, Fan J et al (2005). Развитие сетей внимания: исследование фМРТ с детьми и взрослыми. Нейроизображение 28 : 429–439.
ПабМед Google ученый
Корнак Д.
Р., Ракич П. (1995). Радиальное и горизонтальное размещение клонально родственных клеток в неокортексе приматов: связь с различными митотическими линиями. Нейрон 15 : 311–321.КАС пабмед Google ученый
Костович И., Гольдман-Ракич П.С. (1983). Транзиторное окрашивание холинэстеразы в медиодорсальном ядре таламуса и его связи в развивающемся мозге человека и обезьяны. J Comp Neurol 219 : 431–447.
КАС пабмед Google ученый
Костович И., Иудас М., Петанек З., Ситлич Г. (1995). Онтогенез целенаправленного поведения: анатомо-функциональные соображения. Int J Psychophysiol 19 : 517–527.
Google ученый
Костович И., Иуда М., Радош М., Грабац П. (2002). Ламинарная организация головного мозга плода человека, выявленная с помощью гистохимических маркеров и магнитно-резонансной томографии.
Кора головного мозга 12 : 536–544.ПабМед Google ученый
Костович И., Молливер М.Е. (1974). Новая интерпретация ламинарного развития коры головного мозга: синаптогенез в разных слоях неопаллиума у плода человека. Анат Рек 178 : 395.
Google ученый
Костович И., Ракич П. (1980). Цитология и время возникновения интерстициальных нейронов в белом веществе конечного мозга младенцев и взрослых людей и обезьян. Дж Нейроцитол 9 : 219–242.
КАС пабмед Google ученый
Костович И., Ракич П. (1984). Развитие предстриарных зрительных проекций в головном мозге плода обезьяны и человека, выявленное с помощью временного окрашивания холинэстеразой. J Neurosci 4 : 25–42.
КАС пабмед ПабМед Центральный Google ученый
Костович И.
, Ракич П. (1990). История развития переходной субпластинчатой зоны в зрительной и соматосенсорной коре головного мозга макак и человека. J Comp Neurol 297 : 441–470.КАС пабмед Google ученый
Кремер Х.К., Йесаваж Дж.А., Тейлор Дж.Л., Купфер Д. (2000). Как мы можем узнать о процессах развития из перекрестных исследований или можем? Am J Психиатрия 157 : 163–171.
КАС пабмед Google ученый
Крайс Р., Эрнст Т., Росс Б.Д. (1993). Развитие человеческого мозга: in vivo количественное определение содержания метаболитов и воды с помощью протонной магнитно-резонансной спектроскопии. Magn Reson Med 30 : 424–437.
КАС пабмед Google ученый
Креппнер Дж. М., Раттлер М., Беккет С., Касл Дж., Колверт Э., Грутхьюз С. и др.
(2007). Нормальность и нарушения после глубокой ранней институционной депривации: продольное наблюдение в раннем подростковом возрасте. Dev Psychol 43 : 931–946.ПабМед Google ученый
Курцберг Д., Хилперт П.Л., Кройцер Дж.А., Воган-младший Х.Г. (1984). Дифференциальное созревание корковых слуховых вызванных потенциалов на звуки речи у нормальных доношенных и маловесных детей при рождении. Dev Med Child Neurol 26 : 466–475.
КАС пабмед Google ученый
Квон Х., Рейсс А.Л., Менон В. (2002). Нейронная основа затяжных возрастных изменений зрительно-пространственной рабочей памяти. Proc Natl Acad Sci USA 99 : 13336–13341.
КАС пабмед ПабМед Центральный Google ученый
Ladher R, Schoenwolf GC (2005). Создание нервной трубки: индукция нервной трубки и нейруляция.
В: Махендра С.Р., Якобсон М. (ред.). Нейробиология развития , 4-е изд. Kluwer Academic/Plenum Publishing: Нью-Йорк. стр. 1–20.Google ученый
Landing BH, Shankle WR, Hara J, Brannock J, Fallon JH (2002). Развитие структуры и функций постнатальной коры головного мозга человека от рождения до 72 месяцев: изменение толщины слоев II и III коррелирует с появлением новых возрастных моделей поведения. Педиатр Патол Мол Мед 21 : 321–342.
КАС пабмед Google ученый
Lenroot R, Gogtay N, Greenstein D, Wells E, Wallace G, Clasen L et al (2007). Половой диморфизм траекторий развития мозга в детском и подростковом возрасте. НейроИзображение 36 : 1065–1073.
ПабМед Google ученый
Леунг Х.К., Скудларски П., Гейтенби Дж.К., Петерсон Б.С., Гор Дж.К.
(2000). Связанное с событием функциональное МРТ-исследование задачи интерференции цветовых слов. Кора головного мозга 10 : 552–560.КАС пабмед Google ученый
Леви Д.М., Ли Р.В. (2009). Улучшение работы амблиопии зрительной системы. Philos Trans R Soc Lond B Biol Sci 364 : 399–407.
ПабМед Google ученый
Левитт П. (2003). Структурное и функциональное созревание развивающегося мозга приматов. J Pediatr 143 : S35–S45.
КАС пабмед Google ученый
Лян М., Чжоу И., Цзян Т., Лю З., Тянь Л., Лю Х. и др. (2006). Широко распространенная функциональная разобщенность при шизофрении с функциональной магнитно-резонансной томографией в состоянии покоя. Нейроотчет 17 : 209–213.
ПабМед Google ученый
Lin W, Zhu Q, Gao W, Chen Y, Toh CH, Styner M et al (2008).
Функциональная связность МРТ выявляет корковую функциональную связность в развивающемся мозге. AJNR Am J Neuroradiol 29 : 1883–1889.КАС пабмед ПабМед Центральный Google ученый
Лю Ф., Гарланд М., Дуан Ю., Старк Р.И., Сюй Д., Донг З. и др. (2008). Изучение развития головного мозга плода бабуина с помощью магнитно-резонансной томографии в 3 Тесла. НейроИзображение 40 : 148–159.
ПабМед Google ученый
Лю Ю, Рао М.С. (2004). Глиальные предшественники в ЦНС и возможные родственные отношения между ними. Biol Cell 96 : 279–290.
КАС пабмед Google ученый
Лопес-Бендито Г., Каутинат А., Санчес Х.А., Бьель Ф., Флэймз Н., Гарратт А.Н. и др. (2006). Тангенциальная миграция нейронов контролирует направление аксонов: роль нейрегулина-1 в навигации таламокортикальных аксонов.
Сотовый 125 : 127–142.КАС пабмед ПабМед Центральный Google ученый
Лосси Л., Мериги А. (2003). In vivo клеточные и молекулярные механизмы апоптоза нейронов в ЦНС млекопитающих. Прог Нейробиол 69 : 287–312.
КАС пабмед Google ученый
Лу Л., Леонард С., Томпсон П., Кан Э., Джолли Дж., Добро пожаловать С. и др. (2007). Нормальные изменения развития нижнего лобного серого вещества связаны с улучшением фонологической обработки: продольный анализ МРТ. Кора головного мозга 17 : 1092–1099.
ПабМед Google ученый
Лучана М., Нельсон, Калифорния (1998). Функциональное появление префронтально управляемых систем рабочей памяти у детей от четырех до восьми лет. Нейропсихология 36 : 273–293.
КАС пабмед Google ученый
Луна Б.
, Гарвер К.Е., Урбан Т.А., Лазар Н.А., Суини Дж.А. (2004). Созревание познавательных процессов от позднего детства до взрослой жизни. Ребенок Дев 75 : 1357–1372.ПабМед Google ученый
Luna B, Thulborn KR, Munoz DP, Merriam EP, Garver KE, Minshew NJ et al (2001). Созревание широко распространенных функций мозга способствует когнитивному развитию. Нейроизображение 13 : 786–793.
КАС пабмед Google ученый
Лунд Дж.С., Льюис Д.А. (1993). Нейроны локальной цепи развивающейся и зрелой префронтальной коры макака: Гольджи и иммуноцитохимические характеристики. J Comp Neurol 328 : 282–312.
КАС пабмед Google ученый
Майя Т., Куни Р., Петерсон Б. (2008). Нейронные основы обсессивно-компульсивного расстройства у детей и взрослых.
Развитие психопата 20 : 1251–1283.Google ученый
Малатеста П., Хартфусс Э., Гоц М. (2000). Выделение радиальных глиальных клеток с помощью сортировки флуоресцентно-активированных клеток позволяет выявить происхождение нейронов. Разработка 127 : 5253–5263.
КАС пабмед Google ученый
Марин-Падилья М (1970). Пренатальный и ранний постнатальный онтогенез моторной коры человека: исследование Гольджи. I. Последовательное развитие слоев коры. Мозг Res 23 : 167–183.
КАС пабмед Google ученый
Марин-Падилья М (1971). Ранний пренатальный онтогенез неокортекса коры больших полушарий Felix domestica. Исследование Гольджи. I. Первичная организация неокортекса. Z Anat Entwicklungsgesch 134 : 117–145.
КАС пабмед Google ученый
Марин-Падилья М (1978).
Двойное происхождение неокортекса млекопитающих и эволюция корковой пластинки. Анат Эмбриол 152 : 109–126.КАС Google ученый
Марин-Падилья М. (1988). Ранний онтогенез коры головного мозга человека. В: Питерс А., Джонс Э.Г. (ред.). Развитие и созревание коры головного мозга . Планум: Нью-Йорк. стр. 1–30.
Google ученый
Марш Р., Майя ТВ, Петерсон Б.С. (2009). Функциональные нарушения в лобно-стриарных цепях при множественных детских психопатологиях. Am J Психиатрия 166 : 664–674.
ПабМед ПабМед Центральный Google ученый
Marsh R, Zhu H, Schultz RT, Quackenbush G, Royal J, Skudlarski P et al (2006). Исследование развития саморегулирующегося контроля с помощью фМРТ. Hum Brain Map 27 : 848–863.
ПабМед ПабМед Центральный Google ученый
Martin SJ, Grimwood PD, Morris RG (2000).
Синаптическая пластичность и память: оценка гипотезы. Annu Rev Neurosci 23 : 649–711.КАС пабмед Google ученый
Mata M, Fink DJ, Gainer H, Smith CB, Davidsen L, Savaki H и др. (1980). Зависимый от активности энергетический обмен в задней доле гипофиза крыс в первую очередь отражает активность натриевой помпы. J Нейрохим 34 : 213–215.
КАС пабмед Google ученый
Мацузава Дж., Мацуи М., Кониши Т., Ногучи К., Гур Р.С., Билкер В. и др. (2001). Возрастные объемные изменения серого и белого вещества головного мозга у здоровых младенцев и детей. Кора головного мозга 11 : 335–342.
КАС пабмед Google ученый
Маурер Д., Льюис Т.Л., Брент Х.П., Левин А.В. (1999). Быстрое улучшение остроты зрения у младенцев после визуального ввода.
Наука 286 : 108–110.КАС пабмед Google ученый
Mazoyer B, Zago L, Mellet E, Bricogne S, Etard O, Houde O et al (2001). Корковые сети рабочей памяти и исполнительных функций поддерживают сознательное состояние покоя человека. Brain Res Bull 54 : 287–298.
КАС пабмед Google ученый
McCarthy G, Blamire AM, Puce A, Nobre AC, Bloch G, Hyder F et al (1994). Функциональная магнитно-резонансная томография активации префронтальной коры человека во время задачи на пространственную рабочую память. Proc Natl Acad Sci USA 91 : 8690–8694.
КАС пабмед ПабМед Центральный Google ученый
МакКоннелл С.К., Гош А., Шац С.Дж. (1989). Нейроны подпластины образуют первый аксонный путь от коры головного мозга. Наука 245 : 978–982.
КАС пабмед Google ученый
McGraw P, Liang L, Provenzale JM (2002). Оценка нормальных возрастных изменений анизотропии в младенчестве и детстве по данным диффузионно-тензорной визуализации. AJR Am J Roentgenol 179 : 1515–1522.
ПабМед Google ученый
Макманус М.Ф., Насралла И.М., Гопал П.П., Пэк В.С., Голден Дж.А. (2004). Аксон-опосредованная миграция интернейронов. J Neuropathol Exp Neurol 63 : 932–941.
ПабМед Google ученый
Майнеке Д.Л., Ракич П. (1992). Экспрессия рецепторов ГАМК и ГАМКА нейронами субпластинчатой зоны в развивающейся затылочной коре приматов: свидетельство временных локальных цепей. J Comp Neurol 317 : 91–101.
КАС пабмед Google ученый
Мишель А.
Е., Гари Л.Дж. (1984). Развитие дендритных шипов в зрительной коре человека. Hum Neurobiol 3 : 223–227.КАС пабмед Google ученый
Micheloyannis S, Pachou E, Stam CJ, Breakspear M, Bitsios P, Vourkas M et al (2006). Сети маленького мира и нарушенная функциональная связность при шизофрении. Schizophr Res 87 : 60–66.
ПабМед Google ученый
Миллар В.С., Уотсон Дж.С. (1979). Пересмотрено влияние отсроченной обратной связи на обучение младенцев. Ребенок Дев 50 : 747–751.
КАС пабмед Google ученый
Миллер Р. Х. (2002 г.). Регуляция развития олигодендроцитов в ЦНС позвоночных. Прог Нейробиол 67 : 451–467.
КАС пабмед Google ученый
Мията Т.
, Кавагути А., Окано Х., Огава М. (2001). Асимметричное наследование радиальных глиальных волокон кортикальными нейронами. Нейрон 31 : 727–741.КАС пабмед Google ученый
Молливер М.Е., Костович И., ван дер Лоос Х. (1973). Развитие синапсов в коре головного мозга плода человека. Мозг Res 50 : 403–407.
КАС пабмед Google ученый
Монах К.С., Уэбб С.Дж., Нельсон К.А. (2001). Пренатальное нейробиологическое развитие: молекулярные механизмы и анатомические изменения. Dev Neuropsychol 19 : 211–236.
КАС пабмед Google ученый
Моришита Х., Хенш Т. (2008). Еще раз о критическом периоде: влияние на зрение. Curr Opin Neurobiol 18 : 101–107.
КАС пабмед Google ученый
Моррис Р.
Г. (2006 г.). Элементы нейробиологической теории функции гиппокампа: роль синаптической пластичности, синаптической маркировки и схем. Eur J Neurosci 23 : 2829–2846.КАС пабмед Google ученый
Маунткасл В.Б. (1997). Столбчатая организация неокортекса. Мозг 120 : 701–722.
ПабМед Google ученый
Маунткасл В.Б., Дэвис П.В., Берман А.Л. (1957). Реактивные свойства нейронов соматосенсорной коры кошек на периферические раздражители. J Нейрофизиол 20 : 374–407.
КАС пабмед Google ученый
Мрзляк Л., Уйлингс Х.Б., Костович И., Ван Эден К.Г. (1988). Пренатальное развитие нейронов префронтальной коры человека: I. Качественное исследование Гольджи. J Comp Neurol 271 : 355–386.
КАС пабмед Google ученый
Мрзляк Л.
, Уйлингс Х.Б., Костович И., ван Эден К.Г. (1992). Пренатальное развитие нейронов префронтальной коры человека. II. Количественное исследование Гольджи. J Comp Neurol 316 : 485–496.КАС пабмед Google ученый
Мрзляк Л., Уйлингс Х.Б.М., ван Эден К.Г., Иуда М. (1990). Развитие нейронов в префронтальной коре человека на пренатальной и постнатальной стадиях. В: Uylings HBM, van Eden CG, de Bruin JPC, Corne MA, Feenstra MGP (eds). Префронтальная кора: ее строение, функции и патология . Эльзевир: Амстердам. стр. 185–222.
Google ученый
Мукерджи П., Миллер Дж.Х., Шимони Дж.С., Контуро Т.Е., Ли Б.К., Алмли Ч.Р. и др. (2001). Нормальное созревание мозга в детстве: тенденции развития, характеризуемые диффузионно-тензорной МРТ. Радиология 221 : 349–358.
КАС пабмед Google ученый
Несс Дж.
К., Романко М.Дж., Ротштейн Р.П., Вуд Т.Л., Левисон С.В. (2001). Перинатальная гипоксия-ишемия вызывает апоптотическую и эксайтотоксическую гибель перивентрикулярных олигодендроцитов-предшественников белого вещества. Дев Нейроски 23 : 203–208.КАС пабмед Google ученый
Ньювенхейс Р. (1994). Неокортекс. Анат Эмбриол 190 : 307–337.
КАС Google ученый
Noctor SC, Flint AC, Weissman TA, Dammerman RS, Kriegstein AR (2001). Нейроны, полученные из клеток радиальной глии, образуют радиальные единицы в неокортексе. Природа 409 : 714–720.
КАС пабмед Google ученый
Новак Г.П., Курцберг Д., Кройцер Дж.А., Воган-младший Х.Г. (1989). Корковые реакции на звуки речи и их форманты у нормальных младенцев: последовательность созревания и пространственно-временной анализ.
Электроэнцефалогр Клин Нейрофизиол 73 : 295–305.КАС пабмед Google ученый
Нудо Р.Дж., Мастертон Р.Б. (1986). Индуцированное стимуляцией мечение [14C]2-дезоксиглюкозой синаптической активности в центральной слуховой системе. J Comp Neurol 245 : 553–565.
КАС пабмед Google ученый
О’Доэрти Дж. (2004). Представления о вознаграждении и обучение, связанное с вознаграждением, в человеческом мозгу: выводы из нейровизуализации. Curr Opin Neurobiol 14 : 769–776.
ПабМед Google ученый
О’Доннелл С., Нозеворти, доктор медицины, Левин Б., Деннис М. (2005). Толщина коры лобно-полярной области у типично развивающихся детей и подростков. Нейроизображение 24 : 948–954.
ПабМед Google ученый
О’Рурк Н.
А., Дэйли М.Э., Смит С.Дж., МакКоннелл С.К. (1992). Разнообразные миграционные пути в развивающейся коре головного мозга. Наука 258 : 299–302.ПабМед Google ученый
Олесен П.Дж., Маковяну Дж., Тегнер Дж., Клингберг Т. (2007). Мозговая активность, связанная с рабочей памятью и отвлечением внимания у детей и взрослых. Кора головного мозга 17 : 1047–1054.
ПабМед Google ученый
Олесен П.Дж., Надь З., Вестерберг Х., Клингберг Т. (2003). Комбинированный анализ данных ДТИ и фМРТ выявляет совместное созревание белого и серого вещества в лобно-теменной сети. Brain Res Cogn Brain Res 18 : 48–57.
ПабМед Google ученый
Olson EC, Walsh CA (2002). Гладкое, грубое и перевернутое развитие новой коры. Curr Opin Genet Dev 12 : 320–327.
КАС пабмед Google ученый
Оуэн А.М., Даунс Дж.Дж., Саакян Б.Дж., Полки К.Е., Роббинс Т.В. (1990). Планирование и пространственная рабочая память после поражения лобной доли у человека. Нейропсихология 28 : 1021–1034.
КАС пабмед Google ученый
Партридж С.К., Мукерджи П., Берман Д.И., Генри Р.Г., Миллер С.П., Лу И. и др. (2005). Количественное определение параметров диффузионной тензорной визуализации в трактах белого вещества недоношенных новорожденных на основе трактографии. J Magn Reson Imaging 22 : 467–474.
ПабМед Google ученый
Паус Т. (2005 г.). Картирование созревания мозга и когнитивного развития в подростковом возрасте. Trends Cogn Sci 9 : 60–68.
ПабМед Google ученый
Паус Т.
, Коллинз Д.Л., Эванс А.С., Леонард Г., Пайк Б., Зийденбос А. (2001). Созревание белого вещества головного мозга человека: обзор магнитно-резонансных исследований. Brain Res Bull 54 : 255–266.КАС пабмед Google ученый
Паус Т., Кешаван М., Гидд Дж. (2008). Почему многие психические расстройства возникают в подростковом возрасте? Nat Rev Neurosci 9 : 947–957.
КАС пабмед ПабМед Центральный Google ученый
Пенсеа В., Бингаман К.Д., Фридман Л.Дж., Лускин М.Б. (2001). Нейрогенез в субвентрикулярной зоне и ростральном миграционном потоке переднего мозга новорожденных и взрослых приматов. Опыт Нейрол 172 : 1–16.
КАС пабмед Google ученый
Peterson B, Anderson AW, Ehrenkranz R, Staib LH, Tageldin M, Colson E et al (2003).
Региональные объемы головного мозга и их более поздние корреляты развития нервной системы у доношенных и недоношенных детей. Педиатрия 111 : 939–948.ПабМед Google ученый
Петерсон Б.С., Кейн М.Дж., Александр Г.Э., Лакади С. (2002). Функциональное МРТ-исследование, связанное с событием, сравнивающее интерференционные эффекты в задачах Саймона и Струпа. Cogn Brain Res 13 : 427–440.
Google ученый
Петерсон Б.С., Пайн Д.С., Коэн П., Брук Дж.С. (2001). Проспективное лонгитюдное исследование тиковых, обсессивно-компульсивных и синдрома дефицита внимания/гиперактивности в эпидемиологической выборке. J Am Acad Детская подростковая психиатрия 40 : 685–695.
КАС пабмед Google ученый
Петерсон Б.С., Скудларски П., Гейтенби Дж.К., Чжан Х. (1999).
ФМРТ-исследование интерференции слов-цветов Струпа: доказательства того, что субрегионы поясной извилины обслуживают несколько распределенных систем внимания. Биол Психиатрия 45 : 1237–1258.КАС пабмед Google ученый
Петридес М., Аливисатос Б., Мейер Э., Эванс А.С. (1993). Функциональная активация лобной коры человека при выполнении заданий на вербальную рабочую память. Proc Natl Acad Sci USA 90 : 878–882.
КАС пабмед ПабМед Центральный Google ученый
Пфеффербаум А., Маталон Д.Х., Салливан Э.В., Роулз Дж.М., Ципурский Р.Б., Лим К.О. (1994). Количественное магнитно-резонансное исследование изменений морфологии мозга от младенчества до позднего взросления. Arch Neurol 51 : 874–887.
КАС пабмед Google ученый
Piaget J (1954).
Конструирование реальности у ребенка . Базовый: Нью-Йорк.Google ученый
Пивен Дж., Арндт С., Бейли Дж., Хаверкамп С., Андреасен Н.С., Палмер П. (1995). МРТ-исследование размера мозга при аутизме. Am J Психиатрия 152 : 1145–1149.
КАС пабмед Google ученый
Познер М.И., Петерсен Ю.В. (1990). Система внимания человеческого мозга. Annu Rev Neurosci 13 : 25–42.
КАС пабмед Google ученый
Прехтл Х.Ф., Хопкинс Б. (1986). Эволюционные трансформации спонтанных движений в раннем детстве. Ранний Хум Дев 14 : 233–238.
КАС пабмед Google ученый
Рэдлер Э., Рэдлер А (1978). Авторадиографическое исследование раннего нейрогенеза в неокортексе крыс. Анат Эмбриол 154 : 267–284.
КАС Google ученый
Raichle ME, MacLeod AM, Snyder AZ, Powers WJ, Gusnard DA, Shulman GL (2001). Режим работы мозга по умолчанию. Proc Natl Acad Sci USA 98 : 676–682.
КАС пабмед ПабМед Центральный Google ученый
Ракич П. (1972). Способ миграции клеток в поверхностные слои неокортекса плода обезьяны. J Comp Neurol 145 : 61–83.
КАС пабмед Google ученый
Ракич П. (1978). Миграция нейронов и контактная ориентация в конечном мозге приматов. Postgrad Med J 54 (Приложение 1): 25–40.
ПабМед Google ученый
Ракич П. (1982). Ранние события развития: клеточные линии, приобретение позиций нейронов, а также ареальное и ламинарное развитие. Neurosci Res Program Bull 20 : 439–451.
КАС пабмед Google ученый
Ракич П. (1988). Спецификация областей коры головного мозга. Наука 241 : 170–176.
КАС пабмед Google ученый
Ракич П. (1995). Радиальная против тангенциальной миграции клонов нейронов в развивающейся коре головного мозга. Proc Natl Acad Sci USA 92 : 11323–11327.
КАС пабмед ПабМед Центральный Google ученый
Ракич П. (2003 г.). Эволюционные и эволюционные адаптации кортикальной радиальной глии. Кора головного мозга 13 : 541–549.
ПабМед Google ученый
Ракич П., Буржуа Дж.П., Экенхофф М.Ф., Зечевич Н., Гольдман-Ракич П.С. (1986). Параллельное перепроизводство синапсов в различных областях коры головного мозга приматов.
Наука 232 : 232–235.КАС пабмед Google ученый
Ракич П., Буржуа Дж.П., Гольдман-Ракич П.С. (1994a). Синаптическое развитие коры головного мозга: значение для обучения, памяти и психических заболеваний. Prog Brain Res 102 : 227–243.
КАС пабмед Google ученый
Ракич П., Кэмерон Р.С., Комуро Х. (1994b). Распознавание, адгезия, трансмембранная передача сигналов и подвижность клеток при управляемой миграции нейронов. Curr Opin Neurobiol 4 : 63–69.
КАС пабмед Google ученый
Ракич С., Зечевич Н. (2000). Запрограммированная гибель клеток в развивающемся конечном мозге человека. Eur J Neurosci 12 : 2721–2734.
КАС пабмед Google ученый
Ramanathan R, Wilkemeyer MF, Mittal B, Perides G, Charness ME (1996).
Алкоголь ингибирует межклеточную адгезию, опосредованную человеческим L1. J Cell Biol 133 : 381–390.КАС пабмед Google ученый
Рапопорт Дж., Гогтай Н. (2008). Нейропластичность мозга у здоровых, гиперактивных и психотических детей: данные нейровизуализации. Нейропсихофармакология 33 : 181–197.
ПабМед Google ученый
Раш Б.Г., Гроув Э.А. (2006a). Паттернирование областей и слоев в развивающейся коре головного мозга. Curr Opin Neurobiol 16 : 25–34.
КАС пабмед Google ученый
Раш Б.Г., Гроув Э.А. (2006b). Паттернирование областей и слоев в развивающейся коре головного мозга. Curr Opin Neurobiol 16 : 25–34.
КАС пабмед Google ученый
Ринн М.
, Пикер А., Брэнд М. (2006). Глобальные и локальные механизмы формирования паттернов переднего и среднего мозга. Curr Opin Neurobiol 16 : 5–12.КАС пабмед Google ученый
Рикманн М., Хронвалл Б.М., Вольф Дж.Р. (1977). О развитии непирамидных нейронов и аксонов вне корковой пластинки: ранняя маргинальная зона в виде паллиальной закладки. Анат Эмбриол 151 : 258–307.
Google ученый
Рикманн М., Вольф Дж. Р. (1981). Дифференцировка «предпластинчатых» нейронов в мантии крысы. Bibliotheca Anat 19 : 142–146.
Google ученый
Розенталь А., Жуэ М., Кенврик С. (1992). Аберрантный сплайсинг мРНК молекулы адгезии нервных клеток L1 в семье с Х-сцепленной гидроцефалией. Нат Жене 2 : 107–112.
КАС пабмед Google ученый
Ротбарт М.
К., Познер М.И., Бойлан А. (1990). Регуляторные механизмы в развитии младенцев. В: Эннс Дж. Т. (ред.). Развитие внимания: исследования и теория . Эльзевир/Северная Голландия: Амстардам. стр. 47–66.Google ученый
Рубиа К., Овермейер С., Тейлор Э., Браммер М., Уильямс С.К., Симмонс А. и др. (2000). Функциональная фронтализация с возрастом: картирование траекторий развития нервной системы с помощью фМРТ. Neurosci Biobehav Rev 24 : 13–19.
КАС пабмед Google ученый
Rubia K, Smith AB, Woolley J, Nosarti C, Heyman I, Taylor E et al (2006). Прогрессивное увеличение активации лобно-полосатого мозга от детства к взрослому во время связанных с событиями задач когнитивного контроля. Hum Brain Map 27 : 973–993.
ПабМед ПабМед Центральный Google ученый
Шерф К.
С., Суини Дж.А., Луна Б. (2006). Мозговая основа возрастных изменений зрительно-пространственной рабочей памяти. J Cogn Neurosci 18 : 1045–1058.ПабМед Google ученый
Schmithorst VJ, Wilke M, Dardzinski BJ, Holland SK (2002). Корреляция диффузионной способности и анизотропии белого вещества с возрастом в детстве и подростковом возрасте: исследование поперечного сечения диффузионно-тензорной МРТ. Радиология 222 : 212–218.
ПабМед Google ученый
Schmithorst VJ, Wilke M, Dardzinski BJ, Holland SK (2005). Когнитивные функции коррелируют с архитектурой белого вещества в нормальной детской популяции: диффузионно-тензорное МРТ исследование. Hum Brain Map 26 : 139–147.
ПабМед ПабМед Центральный Google ученый
Шварц М.Л., Ракич П., Гольдман-Ракич П.
С. (1991). Экспрессия раннего фенотипа корковых нейронов: свидетельство того, что подкласс мигрирующих нейронов имеет мозолистые аксоны. Proc Natl Acad Sci USA 88 : 1354–1358.КАС пабмед ПабМед Центральный Google ученый
Selemon LD, Goldman-Rakic PS (1999). Гипотеза редуцированного нейропиля: модель шизофрении на основе схемы. Биол Психиатрия 45 : 17–25.
КАС пабмед Google ученый
Shankle WR, Rafii MS, Landing BH, Fallon JH (1999). Примерное удвоение числа нейронов в постнатальной коре головного мозга человека и в 35 специфических цитоархитектонических областях от рождения до 72 месяцев. Pediatr Dev Pathol 2 : 244–259.
КАС пабмед Google ученый
Shaw P, Eckstrand K, Sharp W, Blumenthal J, Lerch JP, Greenstein D et al (2007).
Синдром дефицита внимания/гиперактивности характеризуется задержкой созревания коры головного мозга. Proc Natl Acad Sci USA 104 : 19649–19654.КАС пабмед ПабМед Центральный Google ученый
Shaw P, Greenstein D, Lerch J, Clasen L, Lenroot R, Gogtay N et al (2006). Интеллектуальные способности и корковое развитие у детей и подростков. Природа 440 : 676–679.
КАС пабмед Google ученый
Shaw P, Kabani NJ, Lerch J, Eckstrand K, Lenroot R, Gogtay N et al (2008). Траектории развития нервной системы коры головного мозга человека. J Neurosci 28 : 3586–3594.
КАС пабмед ПабМед Центральный Google ученый
Снук Л., Полсон Л.А., Рой Д., Филлипс Л., Больё К. (2005). Диффузионно-тензорная визуализация развития нервной системы у детей и молодых людей.
Нейроизображение 26 : 1164–1173.ПабМед Google ученый
Зондереггер П., Ратьен Ф.Г. (1992). Регуляция роста аксонов в нервной системе позвоночных посредством взаимодействия между гликопротеинами, принадлежащими к двум подгруппам суперсемейства иммуноглобулинов. J Cell Biol 119 : 1387–1394.
КАС пабмед Google ученый
Сонуга-Барке Э. (2005 г.). Причинные модели синдрома дефицита внимания/гиперактивности: от обычных простых дефицитов до множественных путей развития. Биол Психиатрия 57 : 1231–1238.
ПабМед Google ученый
Sowell E, Peterson B, Kan E, Woods RP, Yoshii J, Bansal R et al (2007). Половые различия в толщине коры выявлены у 176 здоровых людей в возрасте от 7 до 87 лет. Кора головного мозга 17 : 1550–1560.
ПабМед Google ученый
Соуэлл Э.Р., Делис Д., Стайлз Дж., Джерниган Т.Л. (2001). Улучшение функционирования памяти и созревание лобных долей между детством и юностью: структурное МРТ-исследование. J Int Neuropsychol Soc 7 : 312–322.
КАС пабмед Google ученый
Соуэлл Э.Р., Петерсон Б.С., Томпсон П.М., Добро пожаловать С.Е., Хенкениус А.Л., Тога А.В. (2003a). Картирование корковых изменений на протяжении всей жизни человека. Nat Neurosci 6 : 309–315.
КАС пабмед Google ученый
Соуэлл Э.Р., Томпсон П.М., Холмс С.Дж., Батт Р., Джерниган Т.Л., Тога А.В. (1999). Локализация возрастных изменений в структуре мозга между детством и юностью с помощью статистического параметрического картирования. Нейроизображение 9 : 587–597.
КАС пабмед Google ученый
Соуэлл Э.
Р., Томпсон П.М., Леонард К.М., Добро пожаловать С.Е., Кан Э., Тога А.В. (2004). Продольное картирование толщины коры и роста мозга у нормальных детей. J Neurosci 24 : 8223–8231.КАС пабмед ПабМед Центральный Google ученый
Соуэлл Э.Р., Томпсон П.М., Добро пожаловать С.Э., Хенкениус А.Л., Тога А.В., Петерсон Б.С. (2003b). Корковые аномалии у детей и подростков с синдромом дефицита внимания и гиперактивности. Ланцет 362 : 1699–1707.
ПабМед Google ученый
Копье LP (2000). Мозг подростка и возрастные поведенческие проявления. Neurosci Biobehav Rev 24 : 417–463.
КАС пабмед Google ученый
Squire LR, Stark CE, Clark RE (2004). Медиальная височная доля. Annu Rev Neurosci 27 : 279–306.
КАС пабмед Google ученый
Стейнберг Л.
(2007). Риск принимая в подростковом возрасте новые перспективы из науки о мозге и поведении. Curr Dir Psychol Sci 16 : 55–59.Google ученый
Компакт-диск Stern (2001 г.). Начальное паттернирование центральной нервной системы: сколько организаторов? Nat Rev Neurosci 2 : 92–98.
КАС пабмед Google ученый
Стюарт Г.Р., Перлман А.Л. (1987). Фибронектиноподобная иммунореактивность в развивающейся коре головного мозга. J Neurosci 7 : 3325–3333.
КАС пабмед ПабМед Центральный Google ученый
Супер Х., Сориано Э., Уйлингс Х.Б. (1998). Функции предпластинки в развитии и эволюции неокортекса и гиппокампа. Res Brain Res Rev 27 : 40–64.
ПабМед Google ученый
Swanson HL (1999).
Что развивается в рабочей памяти? Перспектива продолжительности жизни. Dev Psychol 35 : 986–1000.КАС пабмед Google ученый
Такахаши Т., Свобода К., Малинов Р. (2003). Испытайте усиление передачи, управляя рецепторами AMPA в синапсах. Наука 299 : 1585–1588.
КАС пабмед Google ученый
Тамм Л., Менон В., Рейсс А.Л. (2002). Созревание функций мозга связано с реакцией торможения. J Am Acad Детская подростковая психиатрия 41 : 1231–1238.
ПабМед Google ученый
Танг Н., Хе М., О’Риордан М.А., Фаркас К., Бак К., Леммон В. и др. (2006). Этанол ингибирует активацию митоген-активируемых протеинкиназ молекулами адгезии клеток L1. J Нейрохим 96 : 1480–1490.
КАС пабмед ПабМед Центральный Google ученый
Тарабулский Г.
М., Паскуццо К., Мосс Э., Сен-Лоран Д., Бернье А., Сир С. и др. (2008). Вмешательство на основе привязанности для плохо обращающихся семей. Am J Ортопсихиатрия 78 : 322–332.ПабМед Google ученый
Телен К., Кедар В., Паникер А.К., Шмид Р.С., Мидкифф Б.Р., Манесс П.Ф. (2002). Молекула адгезии нервных клеток L1 усиливает интегрин-зависимую миграцию клеток к белкам внеклеточного матрикса. J Neurosci 22 : 4918–4931.
КАС пабмед ПабМед Центральный Google ученый
Томас К.М., Кинг С.В., Франзен, П.Л., Валлийский Т.Ф., Берковиц А.Л., Нолл, округ Колумбия и др. (1999). Развивающее функциональное МРТ-исследование пространственной рабочей памяти. Нейроизображение 10 : 327–338.
КАС пабмед Google ученый
Thompson PM, Sowell ER, Gogtay N, Giedd JN, Vidal CN, Hayashi KM et al (2005).
Структурная МРТ и развитие мозга. Int Rev Neurobiol 67 : 285–323.ПабМед Google ученый
Томпсон П.М., Видал С., Гидд Дж.Н., Гохман П., Блюменталь Дж., Николсон Р. и др. (2001). Картирование изменений мозга в подростковом возрасте выявляет динамическую волну ускоренной потери серого вещества при очень ранней шизофрении. Proc Natl Acad Sci USA 98 : 11650–11655.
КАС пабмед ПабМед Центральный Google ученый
Томсон А.М., Баннистер А.П. (2003). Межламинарные связи в неокортексе. Кора головного мозга 13 : 5–14.
ПабМед Google ученый
Тофт П.Б., Лет Х., Лу Х.К., Придс О., Хенриксен О. (1994). Концентрации метаболитов в развивающемся мозге оценивали с помощью протонной МР-спектроскопии. J Magn Reson Imaging 4 : 674–680.
КАС пабмед Google ученый
Цудзимото С., Ямамото Т., Кавагути Х., Коидзуми Х., Савагути Т. (2004). Активация префронтальной коры, связанная с рабочей памятью у взрослых и детей дошкольного возраста: исследование оптической топографии, связанное с событием. Кора головного мозга 14 : 703–712.
ПабМед Google ученый
Уддин Л.К., Якобони М., Ланге С., Кинан Дж.П. (2007). Самопознание и социальное познание: роль срединных структур коры и зеркальных нейронов. Trends Cogn Sci 11 : 153–157.
ПабМед Google ученый
Ухер Р., Макгаффин П. (2008). Модерация геном переносчика серотонина экологических неблагоприятных факторов в этиологии психических заболеваний: обзор и методологический анализ. Мол Психиатрия 13 : 131–146.
КАС пабмед Google ученый
Ульманн Э.
Дж., Вонг М., Болдуин Р.Л., Бадженару М.Л., Онда Х., Квятковски Д.Дж. и др. (2002). Астроцит-специфические мыши с условным нокаутом TSC1 демонстрируют аномальную организацию нейронов и судороги. Энн Нейрол 52 : 285–296.КАС пабмед Google ученый
ван дер Кнаап М.С., ван дер Гронд Дж., ван Райен П.С., Фабер Дж.А., Валк Дж., Виллемс К. (1990). Возрастные изменения локализованной протонной и фосфорной МР-спектроскопии головного мозга. Радиология 176 : 509–515.
КАС пабмед Google ученый
Ван Эден К.Г., Мрзляк Л., Воорн П., Уйлингс Х.Б. (1989). Пренатальное развитие ГАМКергических нейронов в неокортексе крысы. J Комп Нейрол 289 : 213–227.
КАС пабмед Google ученый
Воган-младший Х.Г., Курцберг Д. (1989). Электрофизиологические показатели нормального и аберрантного созревания коры.
В: Келлавей П., Нобельс Дж. Л. (ред.). Нейрофизиология развития . Издательство Университета Джона Хопкинса: Балтимор, Мэриленд. стр. 263–287.Google ученый
Wager TD, Smith EE (2003). Нейровизуализационные исследования рабочей памяти: метаанализ. Cogn Affect Behav Neurosci 3 : 255–274.
ПабМед Google ученый
Weickert CS, Webster MJ, Colvin SM, Herman MM, Hyde TM, Weinberger DR et al (2000). Локализация рецепторов эпидермального фактора роста и предполагаемых нейробластов в субэпендимальной зоне человека. J Comp Neurol 423 : 359–372.
КАС пабмед Google ученый
Валлийский Дж. А., Виана А. Г., Петрилл С. А., Матиас М. Д. (2007). Вмешательства для международно усыновленных детей и семей: обзор литературы. Социальная служба для детей и подростков J 24 : 285–311.
Google ученый
Вуд Дж.Г., Мартин С., Прайс ДиДжей (1992). Доказательства того, что самые ранние сгенерированные клетки коры головного мозга мышей образуют временную популяцию в субпластинке и маргинальной зоне. Dev Brain Res 66 : 137–141.
КАС Google ученый
Яковлев П.И., Лекур А. (1967). Миелогенные циклы регионального созревания головного мозга. В: Минковский А. (ред.). Региональное развитие мозга в раннем возрасте . Научные публикации Блэквелла: Бостон. стр. 3–70.
Google ученый
Ю Ю.К., Бультье Р.С., Ван С., Ши С.Х. (2009 г.). Специфические синапсы развиваются преимущественно среди сестринских возбуждающих нейронов в неокортексе. Природа 548 : 501–504.
Google ученый
Зечевич Н. (1998).
Синаптогенез в слое I коры головного мозга человека в первой половине гестации. Кора головного мозга 8 : 245–252.КАС пабмед Google ученый
Зечевич Н., Буржуа Дж. П., Ракич П. (1989). Изменения плотности синапсов в моторной коре макак-резусов в течение внутриутробной и постнатальной жизни. Brain Res Dev Brain Res 50 : 11–32.
КАС пабмед Google ученый
Чжай Г., Лин В., Уилбер К.П., Гериг Г., Гилмор Дж. Х. (2003). Сравнения регионарной диффузии белого вещества у здоровых новорожденных и взрослых, выполненные с помощью 3,0-Тл только головного МРТ. Радиология 229 : 673–681.
ПабМед Google ученый
Скачать ссылки
Благодарности
Эта работа частично финансировалась за счет грантов NIMH MH-K0274677 и T32 Mh26434-27.
Информация об авторе
Авторы и организации
Отделение детской и подростковой психиатрии Колумбийского университета и Психиатрического института штата Нью-Йорк, Нью-Йорк, штат Нью-Йорк, США
Gregory Z Tau & Bradley S Peterson
Авторы
- Gregory Z Tau
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Bradley S Peterson
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Автор, ответственный за переписку
Грегори З Тау.
Дополнительная информация
РАСКРЫТИЕ
Авторы заявляют об отсутствии конфликта интересов.
PowerPoint Slide
PowerPoint Slide для рис. 1
Слайд PowerPoint для рис.
2Слайд Powerpoint для фиг. 3
Powerpoint Slide для рис. 4
. Рис. 5
Слайд PowerPoint к рис. 6
Слайд PowerPoint для рис. 7
Права и разрешения
Перепечатка и разрешения
Об этой статье
Выявление паттернов перехода в развитии нервной системы от младенчества к раннему детству и факторов риска, прогнозирующих нисходящий переход
- Такео Като
- Томоко Нисимура
- Нори Такей
Научные отчеты (2022)
Родной язык и среда грамотности и ее связь с социально-экономическим статусом и структурой белого вещества в младенчестве
- Тед К. Турески
- Джозеф Санфилиппо
- Надин Гааб
Структура и функции мозга (2022)
Визуализация мозга плода примата, отличного от человека, с помощью позитронно-эмиссионной томографии (ПЭТ) SV2A
- Саманта Россано
- Такуя Тойонага
- Ричард Э. Карсон
Европейский журнал ядерной медицины и молекулярной визуализации (2022)
Познание: Einflüsse von Essen, Trinken und Bewegung
- Герман Кальхофф
- Кэтрин Синнинген
- Томас Люке
Monatsschrift Kinderheilkunde (2022)
Особенности ЭЭГ-паттерна сонных веретен и его диагностическое значение в онтогенезе
- Ухинов Е. Б.
- Мадаева И.М.
- Колесников С.И.
Вестник экспериментальной биологии и медицины (2022)
Прецизионная психиатрия: таксономия нейронных цепей депрессии и тревоги
1. Cole MW, Bassett DS, Power JD, Braver TS, Petersen SE. Внутренние и вызванные задачами сетевые архитектуры человеческого мозга. Нейрон. 2014; 83: 238–51. [Бесплатная статья PMC] [PubMed] [Google Scholar]
2. Fox MD, Snyder AZ, Vincent JL, Corbetta M, Van Essen DC, Raichle ME. Человеческий мозг внутренне организован в виде динамических, антикоррелированных функциональных сетей. Proc Natl Acad Sci USA. 2005;102:9673–78. [Бесплатная статья PMC] [PubMed] [Google Scholar]
3. Power JD, Cohen AL, Nelson SM, et al. Функциональная сетевая организация человеческого мозга. Нейрон. 2011;72:665–78. [Бесплатная статья PMC] [PubMed] [Google Scholar]
4. Seeley WW, Menon V, Schatzberg AF, et al. Диссоциируемые внутренние сети связи для обработки значимости и исполнительного контроля. Дж. Нейроски. 2007; 27: 2349–56. [Бесплатная статья PMC] [PubMed] [Google Scholar]
5. Insel T, Cuthbert B, Garvey M, et al. Критерии области исследования (RDoC): к новой системе классификации исследований психических расстройств. Am J Психиатрия. 2010; 167:748–51. [PubMed] [Академия Google]
6. Маркофф Дж. Обама стремится ускорить изучение человеческого мозга. http://www.nytimes.com/2013/02/18/science/project-seeks-to-build-map-of-human-brain.html (по состоянию на 18 февраля 2013 г.)
7. Kupfer DJ, Regier Д.А. Неврология, клинические данные и будущее психиатрической классификации в DSM-5. Am J Психиатрия. 2011; 168: 672–74. [PubMed] [Google Scholar]
8. Whiteford HA, Degenhardt L, Rehm J, et al. Глобальное бремя болезней, связанных с психическими расстройствами и расстройствами, связанными с употреблением психоактивных веществ: результаты исследования глобального бремени болезней 2010 года. Lancet. 2013; 382:1575–86. [PubMed] [Академия Google]
9. Гладдинг П.А., Пещера А, Зареян М. и др. Интегрированные терапевтические и диагностические платформы открытого доступа для персонализированной сердечно-сосудистой медицины. J Pers Med. 2013;3:203–37. [Бесплатная статья PMC] [PubMed] [Google Scholar]
10. Buckner RL, Krienen FM, Yeo BT. Возможности и ограничения внутренней функциональной связности МРТ. Нат Нейроски. 2013;16:832–37. [PubMed] [Google Scholar]
11. Коул М.В., Шнайдер В. Сеть когнитивного контроля: интегрированные области коры с диссоциируемыми функциями. Нейроизображение. 2007; 37: 343–60. [PubMed] [Академия Google]
12. Хабер С.Н., Кнутсон Б. Схема вознаграждения: связь анатомии приматов и визуализации человека. Нейропсихофармакология. 2010; 35:4–26. [Бесплатная статья PMC] [PubMed] [Google Scholar]
13. Уильямс Л.М., Дас П., Лидделл Б.Дж., Кемп А.Х., Ренни С.Дж., Гордон Э. Режим функциональной связи в путях миндалевидного тела диссоциирует уровень осведомленности о сигналах страха. Дж. Нейроски. 2006; 26: 9264–71. [Бесплатная статья PMC] [PubMed] [Google Scholar]
14. Kober H, Barrett LF, Joseph J, Bliss-Moreau E, Lindquist K, Wager TD. Функциональная группировка и корково-подкорковые взаимодействия в эмоциях: метаанализ исследований нейровизуализации. Нейроизображение. 2008;42:998–1031. [Бесплатная статья PMC] [PubMed] [Google Scholar]
15. Хугдал К., Райхл М.Е., Митра А., Шпехт К. О существовании обобщенной неспецифической сети, зависящей от задачи. Передний шум нейронов. 2015;9:430. [Бесплатная статья PMC] [PubMed] [Google Scholar]
16. Криттенден Б.М., Митчелл Д.Дж., Дункан Дж. Набор сети в режиме по умолчанию во время требовательного акта исполнительного контроля. электронная жизнь. 2015;4:e06481. [Бесплатная статья PMC] [PubMed] [Google Scholar]
17. Roalf DR, Ruparel K, Gur RE, et al. Нейровизуализирующие предикторы когнитивных функций в стандартизированной батарее нейрокогнитивных функций. Нейропсихология. 2014; 28:161–76. [Бесплатная статья PMC] [PubMed] [Google Scholar]
18. Форстер С., Нуньес Элизальде АО, Касл Э, Бишоп С.Дж. Разгадка тревожного ума: тревога, беспокойство и фронтальная вовлеченность в устойчивое внимание по сравнению с обработкой вне задачи. Кора головного мозга. 2015;25:609–18. [Бесплатная статья PMC] [PubMed] [Google Scholar]
19. Barch DM, Burgess GC, Harms MP и др. Консорциум WU-Minn HCP Функция в человеческом коннектоме: задача-фМРТ и индивидуальные различия в поведении. Нейроизображение. 2013;80:169–89. [Бесплатная статья PMC] [PubMed] [Google Scholar]
20. Ван Эссен Д.К., Смит С.М., Барч Д.М. и др., Консорциум медицинских работников WU-Minn. Проект Human Connectome WU-Minn: обзор. Нейроизображение. 2013; 80: 62–79. [Бесплатная статья PMC] [PubMed] [Google Scholar]
21. Древец В.К., Прайс Дж.Л., Симпсон Дж.Р., младший, и соавт. Субгенуальные аномалии префронтальной коры при расстройствах настроения. Природа. 1997; 386: 824–27. [PubMed] [Google Scholar]
22. Greicius MD, Krasnow B, Reiss AL, Menon V. Функциональная связь в мозге в состоянии покоя: сетевой анализ гипотезы режима по умолчанию. Proc Natl Acad Sci USA. 2003; 100: 253–58. [Бесплатная статья PMC] [PubMed] [Google Scholar]
23. Mulders PC, van Eijndhoven PF, Schene AH, Beckmann CF, Tendolkar I. Функциональная связность в состоянии покоя при большом депрессивном расстройстве: обзор. Neurosci Biobehav Rev. 2015; 56:330–44. [PubMed] [Google Scholar]
24. Хорн А., Оствальд Д., Рейзерт М., Бланкенбург Ф. Структурно-функциональный коннектом и сеть режима по умолчанию человеческого мозга. Нейроизображение. 2014; 102:142–51. [PubMed] [Google Scholar]
25. Sheline YI, Price JL, Yan Z, Mintun MA. Функциональная МРТ в состоянии покоя при депрессии выявляет повышенную связь между сетями через дорсальный нексус. Proc Natl Acad Sci USA. 2010;107:11020–25. [Бесплатная статья PMC] [PubMed] [Google Scholar]
26. Гамильтон Дж.П., Фармер М., Фогельман П., Готлиб И.Х. Депрессивные размышления, сеть режима по умолчанию и темная материя клинической неврологии. Биол психиатрия. 2015;78:224–30. [Бесплатная статья PMC] [PubMed] [Google Scholar]
27. Singh MK, Kesler SR, Hadi Hosseini SM, et al. Аномальные структурные сети серого вещества при большом депрессивном расстройстве. Биол психиатрия. 2013;74:777–85. [Бесплатная статья PMC] [PubMed] [Google Scholar]
28. Коргаонкар М.С., Форнито А., Уильямс Л.М., Грив С.М. Аномальные структурные сети характеризуют большое депрессивное расстройство: анализ коннектома. Биол психиатрия. 2014;76:567–74. [PubMed] [Академия Google]
29. Менон В., Уддин Л.К. Заметность, переключение, внимание и контроль: сетевая модель функции островка. Структура мозга Функц. 2010; 214:655–67. [Бесплатная статья PMC] [PubMed] [Google Scholar]
30. Peterson A, Thome J, Frewen P, Lanius RA. Исследования нейровизуализации в состоянии покоя: новый способ выявления различий и сходств между тревожными расстройствами? Can J Психиатрия. 2014; 59: 294–300. [Бесплатная статья PMC] [PubMed] [Google Scholar]
31. Kaiser RH, Andrews-Hanna JR, Wager TD, Pizzagalli DA. Крупномасштабная сетевая дисфункция при большом депрессивном расстройстве: метаанализ функциональной связи в состоянии покоя. Джама Психиатрия. 2015; 72: 603–11. [Бесплатная статья PMC] [PubMed] [Google Scholar]
32. Робинсон О.Дж., Крымский М., Либерман Л., Аллен П., Витал К., Гриллон С. Дорсально-медиальная префронтальная (передняя поясная кора) цепь аверсивного усиления коры-амигдалы при нелекарственных генерализованных и социальных тревожных расстройствах: обсервационное исследование. Ланцет Психиатрия. 2014; 1: 294–302. [Бесплатная статья PMC] [PubMed] [Google Scholar]
33. Эткин А., Пратер К.Е., Хоефт Ф., Менон В., Шацберг А.Ф. Нарушение активации передней поясной извилины и связи с миндалевидным телом во время имплицитной регуляции обработки эмоций при генерализованном тревожном расстройстве. Am J Психиатрия. 2010;167:545–54. [Бесплатная статья PMC] [PubMed] [Google Scholar]
34. Stuhrmann A, Suslow T, Dannlowski U. Обработка лицевых эмоций при большой депрессии: систематический обзор результатов нейровизуализации. Биол Тревожное расстройство настроения. 2011;1:10. [Бесплатная статья PMC] [PubMed] [Google Scholar]
35. Klumpp H, Post D, Angstadt M, Fitzgerald DA, Phan KL. Реакция передней поясной коры и островка во время косвенной и прямой обработки эмоциональных лиц при генерализованном социальном тревожном расстройстве. Биол Тревожное расстройство настроения. 2013;3:7. [Бесплатная статья PMC] [PubMed] [Google Scholar]
36. Виктор Т.А., Фьюри М.Л., Фромм С.Дж., Охман А., Древец В.К. Взаимосвязь между реакцией миндалевидного тела на лица в масках и настроением и лечением при большом депрессивном расстройстве. Арх генерал психиатрия. 2010;67:1128–38. [Бесплатная статья PMC] [PubMed] [Google Scholar]
37. Fu CH, Williams SC, Cleare AJ, et al. Ослабление нервной реакции на грустные лица при большой депрессии при лечении антидепрессантами: проспективное исследование функциональной магнитно-резонансной томографии, связанное с событиями. Арх генерал психиатрия. 2004; 61: 877–89.. [PubMed] [Google Scholar]
38. Arnone D, McKie S, Elliott R, et al. Повышение реакции миндалевидного тела на грустные, но не полные страха лица при большой депрессии: связь с настроением и фармакологическим лечением. Am J Психиатрия. 2012; 169:841–50. [PubMed] [Google Scholar]
39. Jaworska N, Yang XR, Knott V, Macqueen G. Обзор исследований фМРТ во время визуально-эмоциональной обработки при большом депрессивном расстройстве. Всемирная биологическая психиатрия. 2015; 16: 448–71. [PubMed] [Google Scholar]
40. Fonzo GA, Ramsawh HJ, Flagan TM, et al. Общие и специфичные для расстройств нейронные реакции на эмоциональные лица при генерализованной тревоге, социальной тревоге и панических расстройствах. Бр Дж. Психиатрия. 2015;206:206–15. [Бесплатная статья PMC] [PubMed] [Google Scholar]
41. Штейн М.Б., Голдин П.Р., Сарин Дж., Зоррилла Л.Т., Браун Г.Г. Повышенная активация миндалевидного тела в ответ на сердитые и презрительные лица при генерализованной социальной фобии. Арх генерал психиатрия. 2002; 59: 1027–34. [PubMed] [Google Scholar]
42. Phan KL, Fitzgerald DA, Nathan PJ, Tancer ME. Связь между гиперактивностью миндалевидного тела по отношению к резким лицам и выраженностью социальной тревожности при генерализованной социальной фобии. Биол психиатрия. 2006; 59: 424–29. [PubMed] [Google Scholar]
43. Killgore WD, Britton JC, Schwab ZJ, et al. Корково-лимбические реакции на замаскированные аффективные лица при посттравматическом стрессовом расстройстве, паническом расстройстве и специфической фобии. Подавить тревогу. 2014; 31: 150–59. [Бесплатная статья PMC] [PubMed] [Google Scholar]
44. Blair KS, Geraci M, Smith BW, et al. Снижение активности дорсальной передней части поясной извилины во время эмоциональной регуляции и контроля внимания сверху вниз при генерализованной социальной фобии, генерализованном тревожном расстройстве и коморбидной генерализованной социальной фобии/генерализованном тревожном расстройстве. Биол психиатрия. 2012; 72: 476–82. [Бесплатная статья PMC] [PubMed] [Google Scholar]
45. Matthews SC, Strigo IA, Simmons AN, Yang TT, Paulus MP. Снижение функциональной связи миндалевидного тела и надкостной поясной извилины связано с усилением депрессии у нелекарственных людей с текущим большим депрессивным расстройством. J Аффективное расстройство. 2008; 111:13–20. [PubMed] [Академия Google]
46. Пратер К.Е., Хосанагар А., Клумпп Х., Ангштадт М., Фан К.Л. Аберрантная связность миндалевидного тела и лобной коры во время восприятия испуганных лиц и в состоянии покоя при генерализованном социальном тревожном расстройстве. Подавить тревогу. 2013;30:234–41. [Бесплатная статья PMC] [PubMed] [Google Scholar]
47. Steffens DC, Taylor WD, Denny KL, Bergman SR, Wang L. Структурная целостность крючковидного пучка и функциональная связность вентральной префронтальной коры в состоянии покоя в пожилом возрасте депрессия. ПЛОС Один. 2011;6:e22697. [Бесплатная статья PMC] [PubMed] [Google Scholar]
48. Zhao YJ, Du MY, Huang XQ, et al. Аномалии серого вещества головного мозга у пациентов с большим депрессивным расстройством, не принимающих лекарства: метаанализ. Психомед. 2014;44:2927–37. [PubMed] [Google Scholar]
49. Treadway MT, Zald DH. Переосмысление ангедонии при депрессии: уроки трансляционной нейронауки. Neurosci Biobehav Rev. 2011; 35: 537–55. [Бесплатная статья PMC] [PubMed] [Google Scholar]
50. Kim MJ, Hamilton JP, Gotlib IH. Снижение объема хвостатого серого вещества у женщин с большим депрессивным расстройством. Психиатрия рез. 2008; 164:114–22. [Бесплатная статья PMC] [PubMed] [Google Scholar]
51. Keedwell PA, Andrew C, Williams SC, Brammer MJ, Phillips ML. Нейронные корреляты ангедонии при большом депрессивном расстройстве. Биол психиатрия. 2005; 58: 843–53. [PubMed] [Google Scholar]
52. Дихтер Г.С., Козинк Р.В., МакКлернон Ф.Дж., Смоски М.Дж. Ремитированная большая депрессия характеризуется гиперактивацией сети вознаграждения во время ожидания вознаграждения и гипоактивацией во время результатов вознаграждения. J Аффективное расстройство. 2012; 136:1126–34. [Бесплатная статья PMC] [PubMed] [Google Scholar]
53. Williams LM, Korgaonkar MS, Song YC, et al. Реакция миндалевидного тела на эмоциональные лица в прогнозировании общей и специфической реакции на лечение антидепрессантами в рандомизированном исследовании iSPOT-D. Нейропсихофармакология. 2015;40:2398–408. [Бесплатная статья PMC] [PubMed] [Google Scholar]
54. Blair K, Shaywitz J, Smith BW, et al. Реакция на эмоциональные выражения при генерализованной социальной фобии и генерализованном тревожном расстройстве: свидетельство отдельных расстройств. Am J Психиатрия. 2008;165:1193–202. [Бесплатная статья PMC] [PubMed] [Google Scholar]
55. Ottaviani C, Cevolani D, Nucifora V, et al. Реакции миндалевидного тела на замаскированные и испуганные лица с низкой пространственной частотой: предварительное исследование фМРТ при паническом расстройстве. Психиатрия рез. 2012; 203:159–65. [PubMed] [Google Scholar]
56. Zhang WN, Chang SH, Guo LY, Zhang KL, Wang J. Нейронные корреляты обработки, связанной с вознаграждением, при большом депрессивном расстройстве: метаанализ исследований функциональной магнитно-резонансной томографии. J Аффективное расстройство. 2013; 151: 531–39. [PubMed] [Google Scholar]
57. Форнито А., Харрисон Б.Дж., Залески А., Саймонс Дж.С. Конкурентная и совместная динамика крупномасштабных функциональных сетей мозга, поддерживающих память. Proc Natl Acad Sci USA. 2012; 109:12788–93. [Бесплатная статья PMC] [PubMed] [Google Scholar]
58. Qiu C, Liao W, Ding J, et al. Изменения региональной гомогенности при социальном тревожном расстройстве: исследование фМРТ в состоянии покоя. Психиатрия рез. 2011; 194:47–53. [PubMed] [Google Scholar]
59. Sylvester CM, Corbetta M, Raichle ME, et al. Функциональная сетевая дисфункция при тревоге и тревожных расстройствах. Тренды Нейроси. 2012; 35: 527–35. [Бесплатная статья PMC] [PubMed] [Google Scholar]
60. Ниендам Т.А., Лэрд А.Р., Рэй К.Л., Дин Ю.М., Глан Д. К., Картер К.С. Метааналитические данные о высшей сети когнитивного контроля, обслуживающей различные исполнительные функции. Cogn Affect Behav Neurosci. 2012;12:241–68. [Бесплатная статья PMC] [PubMed] [Google Scholar]
61. Коргаонкар М.С., Грив С.М., Эткин А., Кослоу С.Х., Уильямс Л.М. Использование стандартизированных протоколов фМРТ для выявления паттернов нарушения регуляции префронтальной цепи, которые являются общими и специфическими для когнитивных и эмоциональных задач при большом депрессивном расстройстве: результаты первой волны исследования iSPOT-D. Нейропсихофармакология. 2013; 38: 863–71. [Бесплатная статья PMC] [PubMed] [Google Scholar]
62. Зигл Г.Дж., Томпсон В., Картер К.С., Штайнхауэр С.Р., Тазе М.Э. Увеличение миндалевидного тела и снижение дорсолатеральных префронтальных ответов BOLD при униполярной депрессии: связанные и независимые признаки. Биол психиатрия. 2007; 61: 198–209. [PubMed] [Google Scholar]
63. Aizenstein HJ, Butters MA, Wu M, et al. Измененное функционирование схемы исполнительного контроля при депрессии позднего возраста: эпизодические и стойкие явления. Am J Гериатр Психиатрия. 2009; 17:30–42. [Бесплатная статья PMC] [PubMed] [Google Scholar]
64. Бартова Л., Мейер Б.М., Диерс К. и соавт. Уменьшено подавление сети в режиме по умолчанию во время задачи на рабочую память при ремиссии большой депрессии. J Psychiatr Res. 2015;64:9–18. [Бесплатная статья PMC] [PubMed] [Google Scholar]
65. Zhu X, Wang X, Xiao J, et al. Доказательства паттерна диссоциации в сетевом подключении по умолчанию в состоянии покоя у пациентов с первой депрессией, ранее не получавших лечения. Биол психиатрия. 2012;71:611–17. [PubMed] [Google Scholar]
66. Арнольд Антерапер С., Триантафиллоу С., Сойер А.Т., Хофманн С.Г., Габриэли Д.Д., Уитфилд-Габриэли С. Гиперсвязность подкорковых сетей в состоянии покоя при социальном тревожном расстройстве. Мозговой контакт. 2014; 4:81–90. [PubMed] [Google Scholar]
67. Matsuo K, Glahn DC, Peluso MA, et al. Префронтальная гиперактивация во время задания на рабочую память у нелеченных людей с большим депрессивным расстройством. Мол Психиатрия. 2007; 12:158–66. [PubMed] [Google Scholar]
68. Saveanu R, Etkin A, Duchemin AM, et al. Международное исследование по прогнозированию оптимального лечения депрессии (iSPOT-D): результаты острой фазы лечения антидепрессантами. J Psychiatr Res. 2015; 61:1–12. [PubMed] [Академия Google]
69. Данлоп Б.В., Майберг Х.С. Биомаркеры на основе нейровизуализации для выбора лечения большого депрессивного расстройства. Диалоги Clin Neurosci. 2014; 16: 479–90. [Бесплатная статья PMC] [PubMed] [Google Scholar]
70. Chen CH, Suckling J, Ooi C, et al. Функциональная связь миндалевидного тела у пациентов с депрессией, получающих лечение антидепрессантами. Нейропсихофармакология. 2008; 33: 1909–18. [PubMed] [Google Scholar]
71. Dunlop BW, Nemeroff CB. Роль дофамина в патофизиологии депрессии. Арх генерал психиатрия. 2007; 64: 327–37. [PubMed] [Академия Google]
72. Biswal BB, Mennes M, Zuo XN, et al. К открытию науки о функции человеческого мозга. Proc Natl Acad Sci USA. 2010; 107:4734–39. [Бесплатная статья PMC] [PubMed] [Google Scholar]
73. Siegle GJ. Комментарий по ту сторону депрессии: зачем ты, депрессивная клиника завтрашнего дня? Clin Psychol (Нью-Йорк) 2011; 18: 305–10. [Бесплатная статья PMC] [PubMed] [Google Scholar]
74. Dichter GS, Gibbs D, Smoski MJ. Систематический обзор отношений между функциональной МРТ в состоянии покоя и реакцией на лечение при большом депрессивном расстройстве. J Аффективное расстройство. 2014; 172С:8–17. [Бесплатная статья PMC] [PubMed] [Google Scholar]
75. Hilimire MR, Mayberg HS, Holtzheimer PE, et al. Влияние глубокой стимуляции подмозолистого пояса на негативную самооценку у пациентов с резистентной к лечению депрессией. Стимулятор мозга. 2015; 8: 185–91. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Оптимизация с помощью Python — примеры моделирования Jupyter Notebook
HomeResourcesОптимизация с помощью Python — примеры моделирования Jupyter Notebook
Примеры моделирования ноутбуков Jupyter
Введение в примеры моделирования Gurobi Jupyter Notebook
Примеры моделирования Gurobi Jupyter Notebook — это модели математической оптимизации, закодированные с использованием Gurobi Python API и реализованные с помощью Jupyter Notebooks.
Эти примеры моделирования Jupyter Notebook:
- Научат вас строить математические модели оптимизации реальных деловых, инженерных или научных задач с помощью Python.
- Проиллюстрируйте широкую применимость математической оптимизации в различных отраслях.
- Объясните важные функции объектов моделирования Gurobi Python API, такие как добавление переменных решения, ограничений и целевой функции, а также построение линейных выражений.
Примеры моделирования Gurobi Jupyter Notebook основаны на реальных примерах использования и проблемах, обсуждаемых в пятом издании книги «Построение моделей в математическом программировании» Х. Пола Уильямса — отличного образовательного ресурса для тех, кто интересуется моделированием. Наша подборка содержит примеры из самых разных отраслей, включая производство, финансовые услуги, энергетику и коммунальные услуги, телекоммуникации и нефть и газ, а также примеры, которые имеют отношение к различным отраслям, таким как оптимизация маркетинговых кампаний и проектирование сети поставок.
Независимо от того, являетесь ли вы начинающим, средним или продвинутым пользователем, эти примеры моделирования Gurobi Jupyter Notebook помогут вам улучшить свои навыки моделирования оптимизации с использованием Python.
Доступ к примерам через Google Colab или GitHub
Вы можете получить доступ к каждому примеру моделирования Jupyter Notebook в Google Colab по ссылкам ниже, если не указано иное. Google Colab – это бесплатная онлайн-среда Jupyter Notebook, которая позволяет писать и выполнять код Python через браузер.
Вы также можете найти примеры Jupyter Notebook на GitHub. Всю коллекцию примеров можно скачать по этой ссылке.
Если вам нужны примеры для наших интерфейсов C, C++, C#, Java, Visual Basic и Python, MATLAB® и R, дистрибутив Gurobi включает обширный набор примеров, иллюстрирующих часто используемые функции библиотек Gurobi. Мы рекомендуем вам начать с чтения обзора примеров (доступно здесь). Однако, если вы хотите сразу перейти к конкретным примерам, посетите нашу страницу примеров функционального кода здесь.
Изучите наш список примеров, чтобы найти уровень, который подходит именно вам фон в дисциплине, которая использует количественные методы.
| Пример | Описание |
|---|---|
| Введение в математическое оптимизационное моделирование | Изучите ключевые компоненты постановки задач смешанно-целочисленного программирования (MIP). Вы узнаете, как использовать оптимизатор Gurobi для вычисления оптимального решения модели MIP. |
Примеры моделирования
*Задачи из пятого издания книги «Построение моделей в математическом программировании» Х. Пола Уильямса.
Новичок
Эти примеры моделирования предназначены для начального уровня, когда мы предполагаем, что вы знаете Python и имеете некоторые знания о построении математических моделей оптимизации.
| Пример | Описание | Промышленность |
|---|---|---|
| Создание оптимального состава для фэнтези-баскетбола | Использует машинное обучение с учителем для прогнозирования результативности баскетболистов на основе исторических данных и формулирует модель целочисленного программирования для создания оптимального состава | Фэнтези-спорт |
| 3D Крестики-нолики* | Расставьте крестики и нолики на трехмерной доске «Крестики-нолики», чтобы свести к минимуму количество завершенных линий или диагоналей. | Общий |
| Покрытие вышек сотовой связи | Простая задача покрытия, при которой строится сеть вышек сотовой связи для обеспечения покрытия сигналом максимально возможного числа людей. | Телекоммуникации |
| Изогнутый фитинг* | Задача подбора линейной функции к набору наблюдений формулируется как задача линейного программирования. | Общий |
| Диетическая проблема | Использование стандартной задачи о диете для демонстрации пакета ticdat**. | Общий |
| Местоположение объекта | Проблема расположения объекта, которая включает строительство складов для снабжения определенного количества супермаркетов. | Логистика |
| Оптимизация маркетинговой кампании | Проблема оптимизации маркетинговой кампании, распространенная в сфере банковских и финансовых услуг. | Финансовые услуги |
| Разделение рынка* | Распределите розничных продавцов по двум подразделениям компании, чтобы оптимизировать компромиссы между несколькими целями разделения рынка. | Логистика |
| Оффшорная ветроэнергетика | Свести к минимуму затраты на прокладку подводных кабелей, собирающих электроэнергию, вырабатываемую оффшорной ветровой электростанцией. | Энергетика и коммунальные услуги |
| Проектирование сети снабжения* | Определите, как удовлетворить покупательский спрос при минимальных затратах на доставку. | Логистика |
| COVID-19 Оптимизация пропускной способности объекта* | Эта задача оптимизации пропускной способности медицинского учреждения COVID-19 показывает, как определить оптимальное расположение и вместимость временных медицинских учреждений. | Здравоохранение |
Промежуточный уровень
Эти примеры моделирования относятся к среднему уровню, и мы предполагаем, что у вас есть некоторые знания о построении математических оптимизационных моделей. Кроме того, вы должны знать Python и быть знакомы с Gurobi Python API.
| Пример | Описание | Промышленность |
|---|---|---|
| Ценообразование в сельском хозяйстве* | Определить цены и спрос на молочные продукты в стране, чтобы максимизировать общий доход от продажи этих продуктов | Продовольствие и сельское хозяйство |
| Выбор лучших признаков для прогнозирования | Задача линейной регрессии, которая минимизирует остаточную сумму квадратов при условии, что количество ненулевых весов признаков должно быть меньше или равно заданному верхнему пределу. | Исследования, аналитика и оптимизация |
| Назначение клиента | Рассмотрите оптимальное размещение объектов (из набора возможных местоположений), чтобы свести к минимуму расстояние между объектами компании и ее клиентами. | Логистика |
| Производство электроэнергии* | Проблема выработки электроэнергии (также известная как задача об обязательности установки) путем выбора оптимального набора электростанций для включения, чтобы удовлетворить ожидаемый спрос на электроэнергию в течение 24-часового временного горизонта. | Энергетика и коммунальные услуги |
| Заводское планирование* | Решите задачу планирования производства и создайте оптимальный производственный план для максимизации прибыли. | Производство |
| Производство пищевых продуктов* | Решите задачу оптимизации смешивания с несколькими сырьевыми материалами, объединенными таким образом, чтобы соответствовать установленным ограничениям для наименьшей стоимости. | Производство |
| Логический дизайн* | Задача построения схемы с использованием минимального количества логических элементов ИЛИ-НЕ, которые будут выполнять логическую функцию, заданную таблицей истинности. | Производство |
| Горнодобывающая промышленность* | Решите задачу многопериодного планирования добычи, чтобы оптимизировать добычу на нескольких шахтах за пятилетний период. | Металлургия и горнодобывающая промышленность |
| Открытые горные работы* | Определите, какие места раскопок выбрать, чтобы максимизировать валовую прибыль от добычи руды. | Металлургия и горнодобывающая промышленность |
| НПЗ* | Проблема планирования производства, когда необходимо принять решение относительно того, какие продукты производить и какие ресурсы использовать для производства этих продуктов. | Нефть и газ |
| Проблема маршрутизации и планирования для техников | Узнайте, как сформулировать и решить задачу маршрутизации транспортных средств с несколькими депо с временными окнами. | Транспорт |
| Оптимизация аренды автомобиля* | В этой задаче об аренде автомобилей вы хотите определить, сколько автомобилей нужно владеть, где они должны располагаться и где увеличить ремонтные мощности, чтобы максимизировать прибыль. | Услуги |
| Экономическое планирование* | Это пример проблемы экономического планирования, с которой может столкнуться страна. Цель состоит в том, чтобы определить различные возможные модели роста экономики. Эта задача формулируется как задача линейного программирования с использованием Gurobi Python API и решается с помощью Gurobi Optimizer. | Правительство |
| Анализ эффективности* | Пример анализа эффективности — это задача линейного программирования, решаемая с помощью Gurobi Python API. | Все |
| Проект сети доставки дронами | Применение задачи «Расположение объекта» для проектирования сети дронов, доставляющих автоматические внешние дефибрилляторы в ответ на внебольничную остановку сердца | Здравоохранение |
Продвинутый
Эти примеры моделирования относятся к продвинутому уровню, где мы предполагаем, что вы знаете Python и Gurobi Python API, а также обладаете продвинутыми знаниями в области построения математических моделей оптимизации. Как правило, целевая функция и/или ограничения в этих примерах сложны или требуют расширенных функций Gurobi Python API.
| Пример | Описание | Промышленность |
|---|---|---|
| Планирование децентрализации* | Для заданного набора отделов компании и возможных городов, в которых эти отделы могут быть расположены, определите «наилучшее» расположение каждого отдела, чтобы максимизировать валовую прибыль. | Логистика |
| Планирование ферм* | Проблема планирования производства, когда необходимо принять решение относительно того, какие продукты производить и какие ресурсы использовать для производства этих продуктов. | Продовольствие и сельское хозяйство |
| Планирование рабочей силы* | Решить проблемы кадрового планирования, когда необходимо сделать выбор в отношении найма, обучения, увольнения и составления расписания персонала. | Производство |
| Стандартный пул | Решите проблему объединения, которая распространена в различных отраслях, включая нефтехимическую, очистку сточных вод, горнодобывающую промышленность, производство продуктов питания и напитков, фармацевтику, сети теплообменников и операции цепочки поставок. | Нефть и газ |
| Коммивояжер | Цель задачи коммивояжера (TSP) — найти кратчайший маршрут, который проходит через каждый город по одному разу и возвращается в первоначальный город. | Исследования, аналитика и оптимизация |
| Планирование рабочей силы | Задача оптимизации планирования рабочей силы, связанная с составлением графиков работы и назначением смен персонала для удовлетворения потребности в ресурсах, меняющейся во времени. | Профессиональные услуги |
| Свертывание белков* | Эта модель является примером задачи о сворачивании белков, сформулированной как задача бинарной оптимизации с использованием Gurobi Python API и решенной с помощью Gurobi Optimizer. | Здравоохранение |
| Оптимизация ограничений* | В этом примере мы рассматриваем ограничение модели целочисленного программирования, в которой все переменные решения в ограничении являются бинарными, цель состоит в том, чтобы найти другое ограничение, которое логически эквивалентно исходному ограничению, но это имеет наименьшее возможное абсолютное значение правой части. | Общий |
| Сравнение белков* | Это пример задачи сравнения белков, сформулированной как задача квадратичного назначения с использованием Gurobi Python API и решенной с помощью Gurobi Optimizer. | Здравоохранение |
| Выдача потерянного багажа* | Это пример задачи маршрутизации транспортных средств, сформулированной как задача бинарной оптимизации с использованием Gurobi Python API. | Транспорт |
| Сбор молока* | Это пример задачи маршрутизации транспортных средств с вместимостью, которая сформулирована как задача бинарной оптимизации с использованием Gurobi Python API и решена с помощью Gurobi Optimizer 9.4019 | Транспорт |
| Управление урожайностью* | Это пример задачи управления доходностью, сформулированной как задача стохастического программирования с тремя периодами с использованием Gurobi Python API. | Авиакомпания |
| Оптимизация выбора портфеля | В этом примере мы хотим найти долю портфеля для инвестирования среди набора акций, который уравновешивает риск и доходность. | Финансовые услуги |
Дополнительные пакеты Python:
**ticdat — это пакет Python с открытым исходным кодом, который может упростить работу по кодированию, необходимую разработчикам MIP, чтобы преодолеть разрыв между программированием для проверки концепции и безопасным, хорошо организованным производством. код класса.
Пример/шаблон The Diet Problem ticdat представляет пакет и охватывает:
- распространение работы в версии пакета
- автоматизация процессов тестирования с помощью набора тестов на основе модульных тестов
- и защита математических движков от реальных проблем с целостностью данных
Коммерческая лицензия
Коммерческие пользователи: Gurobi позволяет вам попробовать бесплатную полнофункциональную коммерческую пробную лицензию на 30 дней. За это время вы также получите:
- Бесплатные услуги сравнительного анализа
- Бесплатные услуги по настройке моделей
- Доступ к первоклассной технической поддержке Gurobi
- Два бесплатных часа индивидуальных консультационных услуг
Не можете просмотреть форму? Пожалуйста, нажмите здесь, чтобы открыть его в новом окне.
Примечание для академических пользователей: Академические пользователи в признанных учебных заведениях должны вместо этого получить бесплатную академическую лицензию, а не коммерческую оценочную лицензию.
Оценочные лицензии зарезервированы для наших коммерческих пользователей, которые сначала хотят убедиться, что это соответствует их потребностям.
Студенты, преподаватели и исследователи могут быстро получить бесплатную академическую лицензию.
Вы можете получить бесплатную лицензию и узнать о нашей академической программе здесь.
Спасибо! Информация успешно отправлена.
публикаций | Лаборатория нейронных цепей
2022
Кеткар М., Гюр Б., Молина-Обандо С., Иоанниду М., Мартелли С., Силиес М. (2022)
Зрительные интернейроны первого порядка распределяют отчетливую информацию о контрасте и яркости по путям включения и выключения для достижения стабильного поведения
eLife 11:e74937. doi: 10.7554/eLife.74937
Henning M, Ramos-Traslosheros G, Gür B, Silies M (2022)
8:3. doi: 10.1126/sciadv.abi7112
2021
Ramos-Traslosheros G and Silies M (2021)
Физиологическая основа противопоставления контраста при вычислении движения у дрозофилы
Nature Communications 12: 4987. doi: 10.1038/s41467-021-9062-24903-wtar J, Renaud Y, Albrecht S, Ghavi-Helm Y, Roignant JY, Silies M, Junion G. (2021)
m 6 Метилирование РНК регулирует проксимальную от промотора паузу РНК-полимеразы II
Molecular Cell , 81 : 3356-3367. doi: 10.1016/j.molcel.2021.06.023
Brandão S C, Silies M, Martelli C (2021)
Адаптивная временная обработка запаховых раздражителей
Cell Tissue Research , 383:125-141. doi: 10.1007/s00441-020-03400-9
2020
Кеткар М.Д., Спорар К., Гюр Б., Рамос-Траслошерос Г., Зайферт М., Силиес М. (2020)
Актуальная биология , 30: 657-669, doi: 10.1016/j.cub.2019.12.038
Neuert H, Deing P, Krukkert K, Naffin E, Steffes G, Risse B, Silies M, Klämbt C (2020)
Сигналы Fas2 гомолога NCAM дрозофилы не зависят адгезии.
Разработка , 147(2): pii: dev.181479. дои: 10.1242/dev.181479.
Gür B, Sporar K, Lopez-Behling A, Silies M (2020)
Отчетливая экспрессия калиевых каналов регулирует свойства зрительного ответа нейронов пластинки Drosophila melanogaster.
J Comp Phys A , 206: 273-287, doi: 10.1007/s00359-019-01385-7
2019
Molina-Obando S, Vargas-Fique JF, Schladt M, Gür Ахтар Дж., Бергер Т.К., Силиес М. (2019) Селективность ON в зрении Drosophila представляет собой мультисинаптический процесс, включающий как глутаматергическое, так и ГАМКергическое ингибирование.
eLife , pii: e49373. doi: 10.7554/eLife.49373.
Ахтар Дж., Море П., Альбрехт С., Марини Ф., Кайзер В., Кулкарни А., Войновски Л., Фонтейн Дж.Ф., Андраде-Наварро М.А., Силиес М., Бергер К. (2019)
TAF-ChIP: подход со сверхнизкими затратами для полногеномного анализа иммунопреципитации хроматина.
Life Sci Alliance : e201
2018
Liou NF, Lin SH, Chen YJ, Tsai KT, Yang CJ, Lin TY, Wu TH, Lin HJ, Chen YT, Gohl DM, Silies M, Chou YH (2018 )
Различные популяции локальных интернейронов интегрируются в обонятельную цепь взрослых дрозофил
Nat Communication 9: 2232. doi: 10.1038/s41467-018-04675-x
Пэн Дж., Сантьяго И.Дж., Ан С., Гур Б., Цуй С.К., Су З., Сюй С., Караханян А., Силиес М., Пекот М.Ю. (2018)
Дрозофила Fezf координирует ламинарно-специфическую связь через клеточно-внутреннюю и клеточную -внешние механизмы. eLife 7: e33962. doi: 10.7554/eLife.33962.
Burgos A, Honjo K, Ohyama T, Qian CS, Shin GJ, Gohl DM, Silies M, Tracey WD, Zlatic M, Cardona A, Grueber WB (2018)
Ноцицептивные интернейроны контролируют двигательные пути модуляторов, способствуя побегу у дрозофилы
eLife 7: e26016, doi: 10.7554/eLife.26016.
Ramos-Traslosheros G, Henning M and Silies M (2018)
Motion detection: cells, circuits, algorithms
Neuroforum 24: doi: 10.1515/nf-2017-A028
2017
Neuert H, Yuva Aydemir Y, Silies M # and Klämbt C # (2017)
Различные способы активации APC/C контролируют рост и нейрон-глиальные взаимодействия в развивающемся глазу дрозофилы
Разработка , 144: 4673-4683. # соответствующих авторов
2015
Fisher YE*, Leong JCS*, Sporar K, Ketkar MD, Gohl DM, Clandinin TR and Silies M (2015)
Требуется класс зрительных нейронов с широкопольными свойствами Для локального обнаружения движения
Current Biology , 25: 3178-3189. *равный вклад
Fisher YE*, Silies M* and Clandinin TR (2015)
Избирательность ориентации улучшает обнаружение движения у дрозофилы.
Нейрон , 88: 390-402. * равный вклад
Sugie A, Hakeda-Suzuki S, Suzuki E, Silies M, Shimozono M, Möhl C, Suzuki T, Tavosanis G (2015)
Молекулярное ремоделирование пресинаптической активной зоны фоторецепторов дрозофилы посредством зависимости от активности Обратная связь
Нейрон , 86: 711-725.
2014
Silies M and Clandinin TR (2014)
EM-erging Motion-Detecting Circuits
Текущая биология , 24: R390-392.
Silies M, Gohl DM, and Clandinin TR (2014)
Цепи обнаружения движения у мух: появление в поле зрения
Annual Review of Neuroscience , 37: 307-327.
Gohl DM, Freifeld F, Silies M, Hwa JJ, Horowitz M и Clandinin TR (2014)
Крупномасштабное картирование мест введения мобильных элементов с использованием цифрового кодирования идентичности образца
Genetics , 196: 615-623 .
Кларк Д.А., Фитцджеральд Дж.Е., Алес Дж.М., Силиес М., Гол Д.М., Норсия А.М. и Кладинин Т.Р. (2014)
Мухи и люди используют общую вычислительную стратегию, которая использует статистику естественной сцены для оценки движения
Nature Neuroscience , 17: 296-303.
2013
Silies M*, Gohl DM*, Fisher YE, Freifeld L, Clark DA and Clandinin TR (2013)
Модульное использование периферийных входных каналов настраивает схему обнаружения движения
Neuron 1, 1-79: 127. *равный вклад
Gao XJ, Potter CJ, Gohl DM, Silies M, Katsov AY, Clandinin TR and Luo L (2013)
Специфическая кинематика и двигательные нейроны для аверсивного хемотаксиса у Drosophila
Current Biology , 23: 1163-1172.
2012
Burke C, Huetteroth W, Krashes M, Gohl DM, Silies M, Certel S, Perisse E, Das G, Owald D and Waddell S (2012)
Многоуровневая передача сигналов вознаграждения через октопамин и дофамин у дрозофилы
Природа , 492: 433-437.
Gohl DM, Silies M и Clandinin TR (2012)
Дрозофила как генетическая модель для исследования движения Vision
In: Создание и разрушение нейронных цепей у дрозофилы. Б.А. Хассан, изд. (Спрингер Сайенс энд Бизнес Медиа, ООО).
Manzo A, Silies M, Gohl DM and Scott K (2012)
Motor neurons controlling fluid ingestion in Drosophila
Proc Natl Acad Sci USA , 109: 6309 – 6312.
2011
Gohl DM, Silies MA, Gao XJ, Bhalerao S, Luongo FJ, Lin CC, Potter CJ и Clandinin TR (2011)
Генетически конвертируемая энхансерная ловушка для направленного комбинаторного рассечения экспрессии генов Patterns
Nature Methods 8: 231-237.
Silies M and Klämbt C (2011)
Адгезия и передача сигналов между нейронами и глиальными клетками у дрозофилы
Curr Opin Neurobiol 21: 11-16.
2010
Silies M and Klämbt C (2010)
Зависимая от APC/C-Fzr/Cdh2 регуляция клеточной адгезии контролирует миграцию глии в PNS дрозофилы
Nature Neuroscience 13: 1357-1364.
Silies M, Yuva-Aydemir Y, Franzdottir SR and Klämbt C (2010)
Имагинальный диск глаза как модель для изучения координации развития нейронов и глии.
Флай (Остин) 4: 71-79.
2009
Stork T, Thomas S, Rodrigues F, Silies M, Naffin E, Wenderdel S and Klämbt C (2009)
Drosophila NeurexinIV стабилизирует взаимодействие нейронов и глии на средней линии ЦНС путем связывания с Wrapper
Разработка 136: 1251-1261.
2008
Stork T, Engelen D, Krudewig A, Silies M, Bainton RJ and Klämbt C (2008)
Организация и функция гематоэнцефалического барьера у дрозофилы
Journal of Neuroscience 7-9 7-9 28: .
2007
Silies M, Edenfeld G, Engelen D, Stork T and Klämbt C (2007)
Развитие периферических глиальных клеток у дрозофилы
Neuron Glia Biol . 3: 35-43.
Silies M, Yuva Y, Engelen D, Aho A, Stork T and Klämbt C (2007)
Миграция глиальных клеток в глазном диске
Journal of Neuroscience 27: 13130-13139.
Изменение мозга и поведения | Центр междисциплинарных исследований никотиновой зависимости
Видение и цели
Появилась новая возможность использовать концепции и инструменты из области когнитивной нейробиологии, чтобы ускорить науку и практику вмешательств, направленных на изменение поведения в отношении здоровья. Благодаря достижениям в области нейровизуализации мы узнаем, где и как в мозгу выполняются процессы самоконтроля. Эта работа указывает на центральную роль нейронной активности в дорсолатеральной префронтальной коре в процессах самоконтроля, которые способствуют здоровому выбору. Кроме того, новые данные показывают, что активность префронтальной коры и цепей когнитивного контроля можно модулировать с помощью различных стратегий, включая тренировку когнитивных упражнений, фармакотерапию, транскраниальную стимуляцию постоянным током и обучение нейробиоуправлению.
Программа изменения мозга и поведения в рамках Центра междисциплинарных исследований никотиновой зависимости преобразует исследования когнитивной нейробиологии в клинические вмешательства для мотивации и поддержания изменения поведения в отношении здоровья. Основная предпосылка заключается в том, что можно повысить способность мозга активировать схемы когнитивного контроля для модуляции процессов самоконтроля, которые способствуют поведению, сопряженному с риском для здоровья, и, в конечном итоге, к плохим результатам для здоровья.
♦ Кэрин Лерман, доктор философии.
♦ Джеймс Луххед, доктор философии.
♦ Джанет Одрейн-Макговерн, доктор философии.
♦ Мэри Фальконе, доктор философии.
♦ Джозеф Кейбл, доктор философии.
♦ Рой Гамильтон, доктор философии.
Описание проекта
Нейронная активность, связанная с рабочей памятью, предсказывает будущие рецидивы курения
Кратковременное воздержание от курения ухудшает когнитивные функции, особенно исполнительную функцию, и это играет роль в рецидиве курения. В этом исследовании изучалось, предсказывает ли активность мозга, связанная с рабочей памятью, последующий рецидив курения сверх стандартных клинических и поведенческих показателей. Восемьдесят курильщиков, обратившихся за лечением, прошли два сеанса функциональной магнитно-резонансной томографии (насыщение курения против 24-часового воздержания) во время выполнения визуальной задачи N-back. Затем последовала краткая консультация и краткосрочная попытка бросить курить. Рецидив в течение первых 7 дней биохимически подтверждался наличием метаболита никотина котинина. Среднее изменение сигнала, зависящее от уровня кислорода в крови (BOLD), было извлечено из априорных областей интереса: двусторонней дорсолатеральной префронтальной коры (DLPFC), медиальной лобной/поясной извилины, задней поясной коры (PCC) и вентромедиальной префронтальной коры. Сигнал от этих областей мозга и дополнительные клинические показатели использовались для моделирования состояния исхода, которое затем подтверждалось с помощью методов повторной выборки. Возврат к курению прогнозировался по усилению симптомов отмены, уменьшению левого DLPFC и увеличению процентного изменения сигнала PCC BOLD (воздержание по сравнению с насыщением от курения). Анализ рабочих характеристик приемника показал 81% площади под кривой с использованием этих предикторов, что является значительным улучшением по сравнению с моделью только с клиническими переменными. Сочетание индуцированного воздержанием снижения активации левой ДЛПФК и снижения супрессии РСС может быть прогностическим маркером неблагоприятного исхода, особенно раннего рецидива курения.
Сопротивление курению и транскраниальная стимуляция постоянным током (tDCS)
Такое поведение, как употребление табака, нездоровое питание и малоподвижный образ жизни, имеют далеко идущие последствия для здоровья. Эти поддающиеся изменению модели поведения являются причиной значительной доли смертей от рака, сердечно-сосудистых заболеваний и диабета и наносят значительный экономический ущерб. Тем не менее, многие виды нездорового поведения очень устойчивы к изменениям, несмотря на широкое знание рисков. Хотя теории изменения поведения были выдвинуты для объяснения стойкости такого поведения, немногие рассматривают нейроповеденческие основы. Эти подходы также не учитывают фундаментальный аспект изменения поведения — способность человека проявлять достаточный самоконтроль, чтобы преодолевать искушения немедленного удовлетворения и/или сохранять внимание к долгосрочным целям. Благодаря достижениям в области нейровизуализации мы узнаем, где и как в мозгу осуществляется самоконтроль над решениями и поведением. Эта работа указывает на центральную роль нейронной активности в дорсолатеральной префронтальной коре (DLPFC) в процессах самоконтроля, которые способствуют здоровому выбору. Кроме того, новые данные показывают, что активность префронтальной коры и цепей когнитивного контроля можно модулировать с помощью неинвазивного и безопасного вмешательства: tDCS. Это внутрисубъектное исследование, подтверждающее концепцию, позволит выяснить, может ли один сеанс tDCS (по сравнению с фиктивной стимуляцией) уменьшить тягу к курению, вызванную стимулом, и повысить способность сопротивляться курению.
СДВГ и транскраниальная стимуляция постоянным током (tDCS) пилот
Синдром дефицита внимания и гиперактивности (СДВГ) характеризуется симптомами импульсивности, невнимательности и гиперактивности, которые появляются в детстве и часто сохраняются во взрослом возрасте. Эти симптомы сопровождаются дефицитом когнитивного контроля и принятием рискованных решений, которые могут привести к негативным психосоциальным последствиям и последствиям для здоровья. Благодаря достижениям в области нейровизуализации мы узнаем, где и как в мозгу осуществляется самоконтроль над решениями и поведением. Эта работа указывает на центральную роль нейронной активности в дорсолатеральной префронтальной коре (DLPFC) в процессах самоконтроля, которые способствуют здоровому выбору. Кроме того, новые данные показывают, что активность префронтальной коры и цепей когнитивного контроля можно модулировать с помощью неинвазивного и безопасного вмешательства: tDCS. В этом межсубъектном исследовании для проверки концепции будет изучено, может ли один сеанс tDCS 1 миллиампер (мА) или 2 мА (по сравнению с фиктивной стимуляцией) улучшить исполнительную когнитивную функцию у взрослых; мы также будем исследовать зависимость доза-реакция, чтобы определить оптимальную дозировку tDCS для улучшения когнитивной функции при СДВГ.
Переподготовка когнитивных функций
Рискованное для здоровья поведение, такое как употребление табака и алкоголя, нездоровое питание и отсутствие физической активности, имеют далеко идущие последствия для профилактики рака. Более 1 из 4 молодых людей сообщают, что употребляли табак в течение последнего месяца. Примерно 42% молодых людей имеют избыточный вес, при этом 16% соответствуют критерию ожирения. Молодой взрослый возраст является критическим периодом для формирования образа жизни, который сохраняется во взрослой жизни. Эти поддающиеся изменению модели поведения являются причиной более 60% смертей от рака и наносят значительный экономический ущерб. Новые работы в области поведенческой экономики и когнитивной нейробиологии предлагают новую стратегию смещения поведенческого выбора в сторону здоровых долгосрочных целей. Предлагаемое исследование будет: (а) оценивать эффективность нейрокогнитивного тренировочного вмешательства (КТ) для улучшения реакции на меры принятия решений, связанные с импульсивным поведением: задержка дисконтирования и чувствительность к риску; и (б) использовать функциональную МРТ, чтобы проверить, влияют ли изменения в нейронной активности на поведение. Сто семьдесят шесть здоровых участников в возрасте от 18 до 35 лет будут случайным образом распределены по одному из двух условий: (а) когнитивные упражнения в Интернете (когнитивный тренинг; CT), предназначенные для улучшения внимания, рабочей памяти, торможения реакции и общую исполнительную функцию или (b) игровые действия в Интернете для контроля когнитивной стимуляции (когнитивная стимуляция; CS). Оба состояния будут подвергнуты фМРТ на исходном уровне (до вмешательства) и после завершения 10-недельного вмешательства, чтобы проверить влияние КТ на принятие решений. Первичные результаты включают изменения в результатах сигнала BOLD и задачах принятия решений (ставка дисконтирования и чувствительность к риску). Промежуточные результаты включают изменения когнитивных функций, таких как внимание, рабочая память и торможение реакции.
Активные гранты
Основанные на нейробиологии вмешательства в поведение, связанное с риском развития рака (01. 08.15 — 30.07.22)
UPENN TCORS: Обмен сообщениями о табачных изделиях в сложной коммуникационной среде (01.09.13 — 31.08.18)
Повторное обучение нейрокогнитивным механизмам поведения, связанного с риском развития рака (01.07.12 — 30.06.16)
Когнитивный тренинг для лечения никотиновой зависимости (09/30/10 — 06/30/15) *Бесплатное продление*
Избранные публикации
Loughead J, Wileyto EP, Valdez JN, Sanborn P, Tang K, Strasser AA, Ruparel K, Ray R, Gur RC, Lerman C. Влияние воздержания на функцию мозга и когнитивные функции у курильщиков различается в зависимости от генотипа COMT. Молекулярная психиатрия , 2009; 14(8):820-826 (PMID1
45; PMCID3896978). Loughead J, Ray R, Wileyto EP, Ruparel K, Sanborn P, Siegel S, Gur RC, Lerman C. Влияние частичного агониста a4b2 варениклина на активность мозга и рабочую память у воздерживающихся курильщиков. Биологическая психиатрия , 2010; 67(8):715-721 (PMID20207347; PMCID в процессе).
Ashare R, Valdez J, Ruparel K, Albelda B, Hopson R, Keefe J, Loughead J, Lerman C. Ассоциация вызванных воздержанием изменений в функции рабочей памяти и генотипа COMT у курильщиков. Психофармакология , 2013; 230(4):653-662 (PMID23828159; PMC3840089).
Ashare RL, Wileyto EP, Ruparel K, Goelz PM, Hopson RD, Valdez JN, Gur RC, Loughead J, Lerman C. Влияние толкапона на рабочую память и активность мозга у воздерживающихся курильщиков: исследование, подтверждающее концепцию. Лекарственная и алкогольная зависимость , 2013; 133(3):852-856 (PMID24095246; PMCID3814299).
Лерман С., Гу Х., Лохед Дж., Рупарел К., Ян Ю., Штейн Э.А. Крупномасштабная связь мозговой сети предсказывает влияние острого воздержания от никотина на тягу и когнитивные функции. JAMA Psychiatry , 2014; 71(5):523-530 (PMID24622915; PMCID4097018).
Loughead J, Wileyto EP, Ruparel K, Falcone M, Hopson R, Gur R, Lerman C. Нейронная активность, связанная с рабочей памятью, предсказывает рецидив курения в будущем. Neuropsychopharmacology , 2015, 40(6):1311-1320 (PMID25469682; PMCID4397393).
Пресс
Philadelphia Inquirer Статья; 2015: Может ли изменение мозга помочь курильщикам бросить курить?
Philadelphia Inquirer Статья; 2015: Новый подход к визуализации мозга может помочь курильщикам бросить курить
Philadelphia Inquirer Статья; 2015: Могут ли игры для мозга изменить поведение в долгосрочной перспективе?
Трансляционная и нейрокомпьютационная оценка дофаминового рецептора 1 частичного агониста для шизофрении> Клинические испытания> Йельская медицина
Фаза I-II
- ИССЛЕДОВАНИЕ HIC#: 2000027506
- Последний В этом исследовании будет проверено, влияет ли CVL-562 (PF-06412562), новое соединение частичного агониста дофамина-1, на нейронные цепи рабочей памяти у пациентов с ранним эпизодом шизофрении. Общая цель состоит в том, чтобы установить нейровизуализирующие биомаркеры семейства дофаминовых рецепторов 1/дофаминовых рецепторов 5 (D1R/D5R) для ускорения разработки агонистов D1R/D5R у людей для лечения когнитивных нарушений, лежащих в основе функциональной инвалидности при шизофрении, ключевого нерешенного клинического заболевания. и забота об общественном здравоохранении.
- Возраст 18 лет — 45 лет
- Genderboth
- Дата начала 11/10/2020
- Конец Дата12/31/2023
Добровольца сейчас
Цель и описание
Это исследование. CVL-562 (PF-06412562), частичный агонист рецептора допамина-1, на нейронный сигнал областей мозга, участвующих в познании, у пациентов с шизофренией и связанными с ней психотическими расстройствами. Основная цель этого исследования — понять цели нейронной цепи этого соединения, поскольку оно связано с улучшением когнитивных функций при шизофрении с использованием задачи пространственной рабочей памяти (SWM). Вторичная цель будет заключаться в количественной оценке дозозависимого воздействия препарата на точность SWM на основе поведенческих данных, собранных во время сканирования, и изучении влияния на функциональную связность.
Все пациенты будут психически стабильными с ранним течением (начало психотических симптомов в течение последних 5 лет) расстройством шизофренического спектра (например, шизофрения, шизоаффективное расстройство или шизофрениформное расстройство) и будут иметь нарушения рабочей памяти (определяемые как показатели ниже среднего при письме).
n-задняя задача батареи PennCNB). В рамках исследования они будут получать перорально определенные дозы CVL-562 (PF-06412562) в случайном порядке с повторными функциональными магнитно-резонансными томографами и когнитивными тестами во время этих посещений. Наиболее распространенными побочными эффектами этого соединения являются тошнота и головная боль. Это многоцентровое исследование, которое требует усилий в общей сложности 4 исследовательских центров (Колумбия, Государственный университет Нью-Йорка, Стоуни-Брук, UPenn и Йельский университет).Критерии приемлемости
Критерии включения :
- Способность дать информированное согласие (согласно результатам собеседования) и добровольное подписанное информированное согласие до выполнения каких-либо процедур, связанных с исследованием
- Желание и способность проводить необходимые для исследования клинические оценки и магнитно-резонансную томографию (МРТ).
- Соответствует критериям шизофрении, шизоаффективного расстройства или шизофрениформного расстройства Диагностического и статистического руководства по психическим расстройствам-5 (DSM-5) на основе структурированного клинического интервью для DSM-5 (SCID-5).
- Быть в пределах 5 лет от начала психоза на основании SCID-5.
- Обращение за лечением и желание принять ограничения на лечение, связанные с исследованием.
- Нарушение рабочей памяти, основанное на показателях ниже среднего (нормативной производительности здорового взрослого человека) при выполнении задания PennCNB letter nback (1).
- Продемонстрируйте преморбидный IQ ≥80 по шкале Penn Reading Assessment (PRA) (2). PRA коррелирует с другими показателями IQ, включая Тест широкого диапазона достижений (WRAT), но он компьютеризирован, базируется в лаборатории соисследователей Рубена С. Гура и Ракель Э. Гур, и его можно кратко администрировать, что позволяет нам уменьшить бремя оценки при уже продолжительном первом посещении.
- Свободно владеть английским языком.
- Быть преимущественно правшой.
- Клинически стабильное лечение в течение как минимум двух месяцев до рандомизации (без посещений отделения неотложной помощи, госпитализаций или текущих суицидальных/убийственных активных мыслей, намерений или планов).
- Стабильный режим приема психотропных препаратов (может не включать психотропные препараты) в течение как минимум 3 недель до исследования и готовность поддерживать неизменный режим во время исследования. При приеме депо-антипсихотических препаратов участники должны иметь стабильную дозировку как минимум для двух последовательных инъекций в качестве самых последних инъекций. Если вы принимаете Invega Trinza, не должно быть планов по изменению дозировки в ходе исследования.
- Для женщин детородного возраста, отсутствие намерения забеременеть в течение периода исследования и согласие на использование надежного метода контроля над рождаемостью (например, внутриматочная спираль (ВМС), гормональная контрацепция, воздержание, презервативы) во время учебный период. Женщинам будет предложено продолжать использовать метод контрацепции в течение 1 месяца после получения последней дозы лекарства. Любой человек, который забеременеет во время исследования, будет немедленно удален, а обсуждение рисков и преимуществ продолжающейся фармакотерапии будет продолжено на чисто клинических основаниях.
Критерии исключения :
- Любое нестабильное медицинское, психиатрическое или неврологическое состояние (включая активные или иным образом выраженные мысли о самоубийстве или убийстве), которое может потребовать неотложного лечения. Активные медицинские состояния, которые являются легкими или хорошо контролируемыми, не являются исключением, если они не влияют на риск для пациента, метаболизм исследуемого препарата или результаты исследования (например, хорошо контролируемый диабет II типа или гипертония) по мнению исследователя.
- В настоящее время лечится любым из следующих препаратов: оланзапином, клозапином, зипрасидоном или азенапином, чтобы избежать заметного воздействия на D1-рецепторы.
- Любое серьезное неврологическое заболевание, черепно-мозговая травма, эпилепсия или тяжелая травма головы в анамнезе, включая сотрясение мозга с потерей сознания > 15 минут, или психохирургическое вмешательство.
- Серьезные заболевания сердца в анамнезе (ишемия, аритмия).
- Любое клинически значимое отклонение от исходных медицинских скрининговых тестов (электрокардиограмма (ЭКГ), общий анализ крови с дифференциалом (CBC), полный метаболический профиль (CMP).
- Гепатит В или С (по отчету или при тестировании) при наличии отклонений от нормы функциональных проб печени
- Вирус иммунодефицита человека (ВИЧ) или синдром приобретенного иммунодефицита (СПИД) (по отчету или по результатам тестирования) из-за когнитивных эффектов ВИЧ и СПИДа.
- Исходная ЭКГ, показывающая удлиненный интервал QTc (>450 для мужчин, >470 для женщин, поправка Framingham(3)), при повторном измерении, показывающем ту же аномалию.
- Текущий эпизод настроения, соответствующий критериям большого депрессивного эпизода, маниакального или гипоманиакального эпизода.
- Электросудорожная терапия (ЭСТ) или лечение нейростимуляцией в течение последних 6 месяцев в анамнезе или планы начать такое лечение во время исследования.
- Текущая история синдрома дефицита внимания/гиперактивности (СДВГ) с фармакологическим лечением.
- Соответствие SCID-5 умеренное или тяжелое расстройство, связанное с употреблением психоактивных веществ, любого вещества, кроме никотина, в течение 3 месяцев до первоначальной оценки. Если употребление психоактивных веществ дает положительный результат при токсикологическом тестировании мочи, участники будут исключены, если не захотят воздерживаться до тех пор, пока моча не станет чистой в течение предлагаемого периода регистрации.
- Положительный токсикологический анализ мочи на любое вещество, кроме марихуаны, при посещениях 3-6.
- Беременность или намерение забеременеть во время исследования.
- Кормящие/кормящие грудью или планирующие это делать во время исследования
- Любой металл в организме (включая медные ВМС) или другие противопоказания к МРТ.
- Тяжелая клаустрофобия, боль в спине, патологическое ожирение или другое состояние, которое может затруднить продолжительный сеанс МРТ или привести к чрезмерным движениям во время сеанса визуализации. Дальтонизм, косоглазие или другие неисправимые проблемы со зрением. Тех, кто носит очки, попросят использовать безопасные для МРТ очки.
- Использование следующих препаратов в течение как минимум 3 недель до исследования или во время исследования: ночные или дневные стимуляторы рецепторов гамма-аминомасляной кислоты-А (ГАМК-А) длительного действия; противосудорожные препараты, за исключением тех, которые показаны для стабилизации настроения, психостимуляторы или медицинский каннабис. Участники могут быть повторно оценены на соответствие требованиям после того, как они не принимали эти лекарства в течение> 3 недель. Участники могут принимать снотворные, не являющиеся ГАМКергическими препаратами, стимуляторы рецепторов ГАМК-А короткого действия (бензодиазепины, небензодиазепины) до и во время исследования.
- Любое изменение типа или дозы психотропных препаратов в течение 3 недель до первоначального визита или во время исследования, чтобы избежать временных последствий изменения режима приема лекарств. Тип лекарства и доза будут тщательно записаны и использованы в качестве ковариации в анализе. Участники могут быть повторно оценены на соответствие требованиям после того, как они будут принимать стабильную дозу лекарств в течение> 3 недель.
- PF-06412562 метаболизируется P450 CYP3A4. Во избежание фармакокинетических взаимодействий лекарства или вещества, которые индуцируют (барбитураты, карбамазепин и др.) или являются умеренно-сильными ингибиторами (кетоконазол и др., грейпфрутовый сок), исключаются, если они используются в течение 3 недель до и во время исследования.
- Активные попытки отказаться от курения, вейпинга или других никотиновых продуктов в течение 3 недель до исследования или во время исследования. Участники могут быть повторно оценены после того, как попытка бросить курить станет стабильной.